摘 要:麻类作物抗逆性强、生物产量高,被认为是潜在的生态工程植物和高碳汇植物。然而,现有关于麻类作物土壤固碳效应的研究存在高度异质性,且部分土壤生态指标缺乏足够的数据进行标准化合并,限制了对其生态功能的全面评估。本研究基于已发表核心试验数据,采用随机效应Meta分析与无Meta分析合成(SWiM)的混合数据分析方法,系统量化了麻类作物对土壤有机碳(SOC)及相关理化性质的影响。结果显示,与常规非麻类作物或休耕裸地相比,麻类作物对SOC总体呈正向效应(lnRR=0.38),且具有极强的情景依赖性,即其碳效应依赖于对土地利用类型、管理强度及土壤背景条件的综合响应。亚组分析表明,不同种类麻类作物固碳能力差异显著,其中多年生麻类作物(罗布麻、苎麻)的固碳潜力远高于一年生麻类作物(黄麻、亚麻和工业大麻等)。麻类作物种植的土壤碳效应存在“生物量输入-农艺扰动”之间的权衡机制:在边际土地或多年生系统中(如主要用于盐碱地等生态修复的罗布麻),低干扰管理促进了根源碳的稳定积累;而在黄麻轮作等常规耕作的一年生系统中,频繁的机械扰动将抵消生物量输入的固碳效益。SWiM分析表明,SOC的增加并非总是伴随着土壤容重的降低或微生物生物量的增加,说明土壤物理结构与生物过程响应的非同步性。研究认为,麻类作物有较强的固碳潜力,其土壤碳效应受到农艺措施等的综合调控,在制订基于麻类作物的固碳与土壤修复策略时,需要重点考虑作物管理强度。
关键词:麻类作物;土壤碳效应;土壤有机碳;Meta分析;SWiM方法;土壤生态指标
在全球气候变化持续加剧与土壤退化持续扩大的背景下,陆地生态系统碳汇的稳定性正面临新的挑战。气候变暖与极端干旱事件的叠加可能显著加剧生态系统碳流失风险,并削弱初级生产力的恢复能力[1-2]。这一趋势导致维持和增强陆地碳库的稳定性成为农业可持续发展与生态安全的重要议题。在农业生态系统中,土壤是最重要的碳储存库之一,其土壤有机碳(Soil Organic Carbon,SOC)的积累与稳定性在全球碳循环及粮食安全保障中发挥着基础性作用[3]。研究[4]表明,通过优化土壤管理措施,农业系统在一定程度上具备提升SOC储量和增强碳汇功能的潜力。然而,长期的常规化农业管理,特别是频繁耕作与作物残体移除,已导致全球农田土壤结构退化、有机质衰减及微生物功能衰退,显著削弱了土壤的碳汇能力[5]。因此,在减少土壤扰动的同时,提高系统生物碳输入强度,被认为是恢复农田SOC的重要路径[3]。
理论上,具有高生物量生产能力和发达根系系统的作物,通过增加地上与地下碳输入,促进深层碳沉积与物理保护机制的形成,进而延长碳在土壤中的周转时间并提升稳定化潜力[6]。多项高水平的生态修复研究证实,具有庞大根系和高生物量生产力的作物(如多年生深根草本和能源作物),能够通过大量的地上凋落物和地下根系周转,将光合作用固定的碳高效、深层地输送至土壤中,在恶劣或退化环境中成功建立起具有极强弹性的稳定碳汇[4,6]。近年来,宏观尺度的碳氮协同管理研究亦强调,在农业生产体系中统筹碳与养分管理,对提升环境效益与管理效率具有重要意义[7]。在此背景下,麻类作物因其生长迅速、生物量高及根系发达等农艺特征,同时在污染修复与可持续农业体系中展现出多功能应用潜力,是应对土壤退化与气候变化的“可持续环境修复工具”[8-9],被认为可能成为增强农田碳汇能力的候选作物之一。
单纯的高生物量输入并不一定会提高土壤有机碳,植物驱动的固碳效应在集约化管理体系下表现出高度的变异性和不确定性[10]。耕作强度、残体管理方式、土壤类型及气候条件等因素,均可能调节碳输入向稳定碳库转化的效率[5]。以往研究探明,土地利用历史、初始土壤碳背景以及农田管理强度等都是引发这些变异的主要因素。在麻类种植体系中,翻耕频率、收获方式及传统沤麻处理等管理措施,可能改变有机质分解速率与团聚体结构稳定性,从而影响其潜在的固碳效果。因此,麻类作物的固碳潜力并非仅由生物量大小决定,而是受制于“生物碳输入强度”与“农艺管理扰动程度”之间的权衡关系,但这一领域仍然缺少量化的分析研究。目前,关于麻类作物对SOC影响的研究多集中于区域性田间试验,结果存在一定差异,缺乏跨区域整合分析以及对管理调节效应的系统量化评估。为弥补这一不足,本研究遵循PRISMA(Preferred Reporting Items for Systematic Reviews and Meta-Analyses)2020声明[11],采用Meta分析与SWiM并行的混合数据整合方法,基于多区域试验数据对麻类种植对SOC的总体影响进行系统评估,并进一步分析耕作与残体管理方式对固碳效应的调节作用。通过识别影响碳稳定化的关键驱动因子,旨在为科学评价麻类作物种植的土壤生态效应及其在农业碳汇管理中的应用潜力提供定量依据。
1 材料与方法
1.1 文献检索与筛选
1.1.1 数据库与时间范围
本研究遵循PRISMA 2020声明开展系统综述与证据合成。文献检索主要基于Web of Science和Scopus两个英文数据库,并以中国知网(CNKI)作为补充中文数据库,以尽可能覆盖中英文相关文献。检索时间范围设定为2010年1月1日至2025年12月31日,检索围绕“麻类作物(bast fiber crops)”及其潜在的土壤生态效应构建,重点关注土壤有机碳(SOC)、土壤容重(BD)和微生物生物量碳(MBC)等指标。
1.1.2 检索词组合
检索策略采用布尔逻辑运算符构建,核心思路是将“麻类作物”与三大生态主题方向进行交叉组合,以系统捕获不同研究情景下的相关证据。麻类作物×土壤/农田生态:旨在获取关于土壤有机碳(SOC)、容重(BD)、团聚体稳定性及土壤健康等方面的研究;麻类作物×微生物/根际:旨在覆盖微生物生物量碳(MBC)、酶活性、根际微生物群落结构等相关文献;麻类作物×碳/修复/重金属:旨在纳入关于碳固持潜力、重金属植物修复(Phytoremediation)及污染土壤改良的研究。
具体的英文检索式示例为:(hemp OR "Cannabis sativa" OR flax OR "Linum usitatissimum" OR ramie OR "Boehmeria nivea" OR kenaf OR "Hibiscus cannabinus" OR jute OR Corchorus OR Apocynum OR "Apocynum venetum"OR sisal OR "Agave sisalana"OR "bast fiber crop*")AND(soil OR cropland OR farmland OR agroecosystem OR rhizosphere OR phytoremediation)。
在CNKI中采用相应的中文关键词组合(如“麻类作物”“工业大麻”“亚麻”“苎麻”ד土壤”“农田生态”“微生物”等)进行检索。对检索结果按题目和摘要进行初筛,剔除明显无关文献后,进入全文评估阶段。
1.2 纳入标准与数据集构建
文献筛选过程严格遵循PRISMA流程(图1)。具体纳入标准如下:
(1)研究对象:研究对象至少包含一种典型麻类作物(如工业大麻、亚麻、苎麻、黄麻、罗布麻、红麻或剑麻等);
(2)试验设计:研究须为田间试验或与真实土壤环境高度相关的控制试验,并设置明确的对照组(如休耕、裸地或非麻类作物种植体系);
(3)数据要求:研究报告SOC、BD或MBC至少一项指标的定量结果,对于纳入Meta分析的SOC指标,需提供处理组与对照组的均值、变异指标(标准差或标准误)及样本量,或提供可换算为上述统计量的数据。
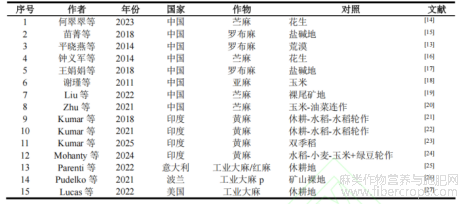
经逐步筛选,从初始检索文献中最终纳入15篇满足定量分析条件的核心试验研究。其中,SOC指标共提取21个对比数据用于Meta分析;BD和MBC可用于定量比较的数据较少(分别为3个和4个对比),仅用于系统评价而不进行数值合并。
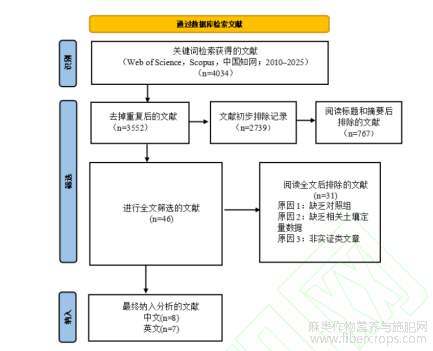
图1 文献筛选PRISMA流程图
1.3 数据提取与效应量计算
从最终纳入的15篇文献中提取作物类型、对照设置、采样深度、试验年限,以及SOC、BD和MBC的数据。
效应量采用自然对数响应比(log response ratio,lnRR)计算:
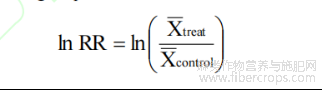
式中:X-treat和X-control分别为麻类作物处理组与对照组的均值;lnRR为无量纲指标。只要同一研究内处理组与对照组的单位一致,即可直接计算并纳入分析。
对于部分仅报告均值而未提供标准差(SD)或标准误(SE)的研究,本研究采用了保守的估算策略。参照Wiebe等[12]的方法,假设变异系数(CV)与同一数据集中的其他相似研究相当,或按照均值的10%进行估算。这种处理方式在保持样本量的同时,尽可能减少了对权重分配的人为干扰。
本研究纳入的大多数文献报告了SOC浓度(g/kg),仅一项研究由平晓燕等[13]报告了SOC储量(g/m2)。鉴于麻类作物在荒漠生态修复中通常具有降低土壤容重(BD)的改良效应,根据碳储量计算公式,在容重降低的情境下,SOC储量的响应比(lnRR)理论上会略低于浓度的响应比。因此,将该储量数据纳入分析属于保守估计,不会人为干扰总体效应量,且能更全面地反映边际土地的修复潜力。
1.4 Meta分析与SWiM综合方法
针对数据标准化程度较高且样本量相对充足的SOC指标,采用随机效应模型开展Meta分析。研究间异质性采用I2统计量进行量化。对于BD和MBC指标,由于可用于定量分析的研究数量有限(k<10),采用SWiM方法进行系统评价,侧重于识别总体响应趋势及潜在不确定性来源。
1.5 发表偏倚与稳健性分析
针对纳入Meta分析的SOC指标,采用漏斗图对效应量分布进行可视化检查,并使用Egger回归检验评估潜在的发表偏倚。鉴于可用研究数量有限,上述检验结果主要用于探索性判断,而非作为严格的否定性数据。
对于采用SWiM综合的BD和MBC指标,由于研究数量有限且数据报告形式高度异质,不适合开展统计意义上的发表偏倚检验。本研究在结果解释与讨论中重点关注数据一致性、研究间差异及不确定性来源,以定性方式评估结论的稳健性。
1.6 统计分析软件
所有Meta分析均在R软件环境(版本4.5.2)中完成,采用metafor等软件包进行效应量计算、模型拟合与结果可视化。
2 结果与分析
2.1 文献筛选与数据集特征
本研究最终纳入于2011–2025年发表的15篇核心试验研究,构成定量与半定量分析的数据基础。研究区域主要分布于中国、印度、美国及欧洲等典型麻类作物种植区,涵盖温带、亚热带及干旱半干旱气候带。在所有纳入文献中,共提取21个SOC对比数据用于Meta分析,由于土壤容重(BD)和微生物生物量碳(MBC)可用于定量合并的研究较少(分别为3个和4个),故仅用于SWiM综合分析。
表1 纳入Meta分析与SWiM综述的研究特征概要
2.2 麻类作物种植对土壤有机碳的总体影响
基于随机效应模型的Meta分析结果(图2),与非麻类作物种植体系对照相比,麻类作物种植总体上显著提升了土壤有机碳(SOC)含量。总体合并效应量(Pooled effect size,lnRR)为0.38(95%CI:0.12~0.63),提升了约46%,且置信区间未跨越零点,表明在统计学上具有显著的正向效应。然而,研究间存在极高的统计学异质性(I²=99.4%,τ²=0.342,p<0.001),说明不同研究中SOC的响应幅度存在巨大差异,需要通过多维度亚组分析来探明其变异来源。

图2 麻类作物种植对土壤有机碳(SOC)总体效应的森林图
2.3 SOC响应的多维度亚组与机制分析
由于总体效应存在极高的异质性,为了深度挖掘SOC响应异质性的来源,打破单一环境变量的局限,本研究从作物生活史、种植制度、农田管理强度、生态区及土地利用类型等5个维度开展了亚组分析。结果显示,麻类作物的固碳效应具有极强的情景依赖性(图3)。

图3 麻类作物种植对土壤有机碳(SOC)影响的多维亚组分析森林图
2.3.1 生物量的输入与农田扰动的权衡机制
涉及农艺管理的三个核心维度上,亚组检验均揭示了极显著或显著的组间差异:多年生与一年生之间(p=0.002)、单作与轮作系统之间(p=0.014)以及低干扰与集约化管理之间(p<0.001)均存在显著的分野。进一步剖析组内具体效应发现,这种差异深刻揭示了麻类作物的净固碳效应严格受制于“生物量碳输入”与“农艺扰动导致的矿化损失”之间的净平衡。
在低干扰系统中,其SOC提升幅度(lnRR=0.88,95%CI:0.35~1.40)呈极显著的正向响应,多年生麻类同样表现出强劲的固碳潜力(lnRR=0.85)。这表明,在缺乏频繁人为耕作干扰的情况下,多年生系统通过持续的根系分泌物输入和细根周转主导了碳固存过程,更重要的是,全周期的少耕或免耕管理最大程度地保护了土壤团聚体,减少了物理保护性有机碳的暴露与分解。正如何翠翠等[14]在我国南方的研究中发现,多年生苎麻显著提高了梯田土壤的SOC,苗菁等[15]和平晓燕等[13]在边际土地上的研究也证实,在缺乏频繁人为耕作干扰的情况下,种植多年生麻类作物能够提升贫瘠土壤的碳库储量。相反,在高强度的常规农业管理下,生物量输入的红利被频繁的扰动大幅削弱甚至彻底抵消。集约化管理(lnRR=0.09)和一年生麻类(lnRR=0.15)的固碳效应虽然为正,但增幅极其微弱。最为典型的证据来自轮作系统,其固碳效应在统计学上已表现为不显著(lnRR=0.06,95%CI:-0.01~0.13,误差线跨越0线)。正如Kumar等[22]的研究,对比了“黄麻-水稻-小麦”轮作与“休耕-水稻-水稻”对照两种种植体系,发现引入黄麻后SOC反而略有下降,这是一年三熟的高强度耕作导致的后果。尽管黄麻提供了大量生物量,但为了播种下一季作物而进行的频繁翻耕加速了土壤呼吸和有机质矿化,导致碳损失速率超过了输入速率。Parenti等[25]关于大麻作为填闲作物的研究也支持了这一观点,频繁的物理破碎会破坏土壤物理保护机制。这从统计学与生态学双重层面表明了缺乏配套的保护性耕作,单纯依靠麻类作物的高生物量根本无法保证农田碳汇的有效增加。
2.3.2 初始碳库基线效应与环境约束
在生态区划与土地利用维度,麻类作物的固碳效应同样展现出受制于环境背景的极显著组间差异。边际土地与常规农田之间(p<0.001)、干旱/半干旱地区与湿润/亚热带地区之间(p=0.042)的响应比截然不同。从组内效应的量级对比来看,边际土地展现出了惊人的固碳跃升(lnRR=1.26,95%CI:0.63~1.89),其效应量是常规农田(lnRR=0.11)的10倍以上;同样,干旱/半干旱地区(lnRR=1.00)的增幅也远超湿润/亚热带地区(lnRR=0.28)。这一悬殊的差异可通过土壤“碳饱和理论”得到解释,即远离碳饱和点的土壤具有极高的固碳效率。
在极度贫瘠的边际土地中,如王娟娟等[17]研究的黄河三角洲裸地,初始SOC仅为0.24g/kg,麻类作物引入后凭借强大的抗逆性与根系开拓能力,迅速填补了巨大的碳饱和亏缺。相比之下,在肥沃的常规农田中,如谢瑾等[18]研究的纳板河流域,初始SOC高达36.83g/kg,碳库已接近饱和。在这种高基线背景下,麻类作物的额外碳投入难以进一步带来显著的SOC跃升,甚至可能引发微生物的“激发效应”,加速原有老碳的分解。这也解释了为何本研究的Meta回归分析(图4)显示种植年限对SOC增量的解释力十分有限(回归斜率p=0.472)。数据的双向印证充分说明麻类作物的土壤固碳潜力并非随时间简单的线性累积,环境背景基线(碳饱和亏缺度)才是决定增量空间的核心要素。这种跨越数量级的基线差异,构成了总体效应极高异质性的根本来源。
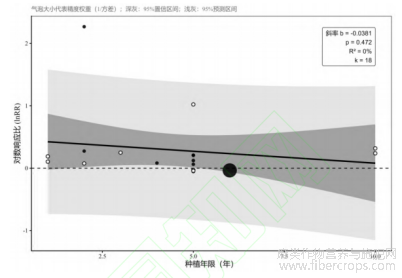
图4 SOC效应量与种植年限关系的Meta回归分析
2.4 土壤容重(BD):SWiM综合分析
基于SWiM综合分析结果,麻类作物种植对土壤物理的核心指标土壤容重(BD)的影响表现出极强的情景依赖性(图5)。值得注意的是,BD作为反向指标,其数值的降低代表着土壤孔隙度的改善。从效应方向与显著性来看,虽然谢瑾等[18]和Kumar等[22]的研究观察到了BD的降低趋势(图5中方块标识),但其95%置信区间均跨越了0线,表明这种物理结构的改善在统计学上并不显著。相反,Mohanty等[24]的最新研究则报告了BD的显著增加(即土壤压实度显著恶化,lnRR>0且未跨越0线)。这一非对称结果表明,单纯种植深根系的麻类作物并不必然带来土壤物理结构的优化。在实际农艺操作中,高强度的田间管理所带来的土壤机械压实效应,极有可能完全覆盖甚至逆转了麻类作物深根系对土壤的生物疏松作用。
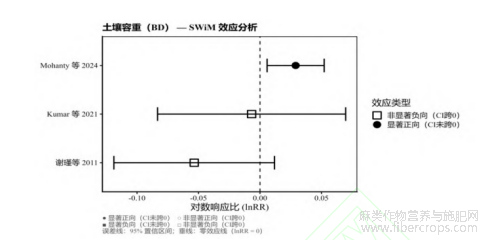
图5 麻类作物对土壤容重(BD)影响的SWiM效应方向图
2.5 土壤微生物量碳(MBC):SWiM综合分析及多维度响应非同步性
作为衡量土壤生物活性的敏感指标,MBC的SWiM综合结果显示出高度不一致性(图6)。在纳入的4个独立观测数据中,呈现出两项显著正向响应(改善,95%CI未跨0线)与两项不显著负向响应(恶化,95%CI跨越0线)并存的复杂态势。结合前文SOC总体效应的显著增加,MBC的这种非同步响应深刻揭示了土壤生态过程的复杂性。例如,Lucas等[27]在美国的田间试验中发现,大麻种植虽然增加了SOC含量,但MBC并没有表现出一致的增加趋势,部分样点甚至出现回落。这反映了土壤微生物群落对环境变化的非线性响应机制,在麻类作物大量且富含难降解木质纤维素的凋落物输入下,土壤微生物群落并非单纯地进行生物量扩增,而是可能正在经历深度的“结构重组”。特定的降解菌群落可能取代了原有的快生型细菌,以适应新的底物特征。这一发现说明,麻类作物对土壤健康的影响是多维且动态的,全面评估其生态效应要摒弃单一的化学指标。本研究采用SWiM分析呈现物理(BD)与生物(MBC)指标的差异化响应,进一步印证了建立多维度评价体系的必要性。
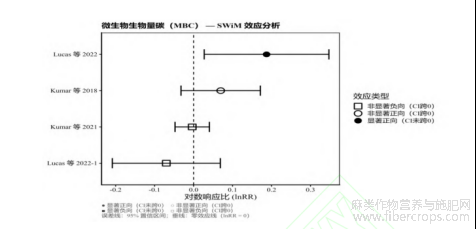
图6 麻类作物种植对微生物生物量碳(MBC)影响的SWiM效应方向图
2.6 发表偏倚与结果稳健性
在系统评价中,识别并处理潜在的偏倚是确保结论可靠性的关键步骤。本研究SOC数据集的漏斗图显示出一定程度的不对称性(图7)。具体而言,散点分布在漏斗图中心线右侧较多,且Egger回归拟合线呈现出明显的倾斜。这提示本研究领域可能存在一定程度的发表偏倚,即报告了麻类作物具有较高固碳效应的小样本研究更容易被发表,而呈现微弱或负向结果的研究可能未被完全检索到。
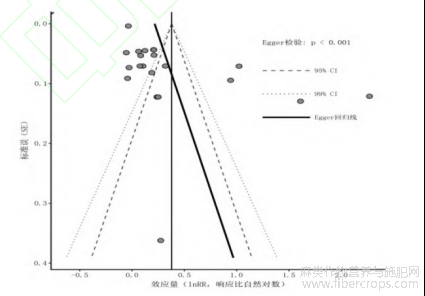
图7 评估SOC发表偏倚的漏斗图
然而,随后的敏感性分析强有力地证实了本研究核心结论的极高稳健性。逐一剔除法的敏感性分析结果表明(图8),在依次剔除任何单项独立研究后,重组的总体效应量(lnRR)仅在0.28至0.40的狭窄区间内波动,始终保持稳定的正向响应,并且所有重组效应量的95%置信区间均稳稳处于0线右侧,未出现跨越0线的情况。这一统计学证据确凿地表明,本研究发现的“麻类作物显著提升农田SOC”的总体正向效应,具有普遍的代表性,并非由某单一极端文献所主导或驱动。
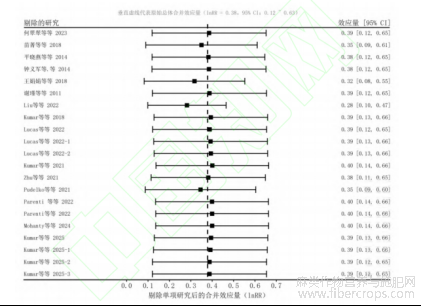
图8 敏感性分析图
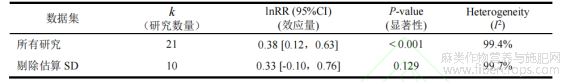
此外,为排除数据标准化过程中(如通过公式估算缺失的标准差SD)可能引入的方差偏差,本研究进行了基于数据质量的敏感性分析(表2)。在剔除11项采用估算SD的研究后,剩余高质量数据集(k=10)的合并效应量为0.33(95%CI:-0.10~0.76)。尽管由于样本量减少导致统计显著性降低(P=0.13),但该效应量的方向和数值与全数据集(lnRR=0.38)高度一致(图2)。这表明估算SD的引入并未改变效应的整体趋势。统计学上的多重稳健性检验,结合多项关于“植物深根系碳输入主导稳定土壤碳库”的高水平生态学文献交叉验证,共同确证了本研究核心发现的可靠性,在适宜的低干扰管理下,麻类作物具备提升土壤有机碳的强大生态潜力。
表2 全数据集与排除估算SD的高质量数据集的Meta分析结果比较
3 讨论
3.1 混合数据分析方法在克服数据稀缺中的优势
本研究最终仅纳入15篇符合严格对照设计与定量数据报告要求的田间研究,反映出当前针对特定作物类型土壤碳效应的高质量实证研究仍然相对有限。类似现象在农业土壤碳研究中并不少见。Poeplau等[28]在对覆盖作物固碳效应进行全球Meta分析时指出,研究间试验周期差异、管理措施不一致以及数据报告不完整,显著限制了可用于效应量合成的样本数量。这说明在农业生态研究中,严格筛选往往会牺牲样本规模,但能够提高结论的可靠性。因此,本研究在数据整合过程中优先保证研究质量,剔除缺乏对照组或关键指标不完整的研究,以降低系统性偏差风险。这种策略符合Meta分析方法学关于研究质量控制与偏倚管理的基本原则[29]。同时,对于难以满足传统效应量计算条件的指标(如BD和MBC),本研究采用方向一致性分析思路进行综合判断。在数据数量有限或研究异质性较高的情况下,通过趋势识别而非强制合并效应量,可以避免统计功效不足导致的误判。这种混合方法论为评估“新兴”或“小众”作物的生态效应提供了可行的分析框架,避免了在数据不足时强行合并所导致的统计错误。
3.2 土地利用强度梯度下的生态效应权衡机制
本研究结果表明,作物种植对SOC的影响具有明显的情景依赖性。这种差异在全球农业土壤研究中已有广泛报道,Lal等[3]指出,土壤碳积累潜力受土地利用历史、初始碳库水平以及管理强度的共同制约。换言之,不同土地生态背景决定了额外碳输入的响应空间。在低碳基线或退化土壤中,增加植被覆盖与根系输入通常有助于SOC恢复;耕作扰动会破坏土壤团聚体结构,使原本受到物理保护的有机碳暴露于微生物分解环境,从而增加矿化风险[2]。这一机制解释了为何在部分常规耕作系统中,高生物量投入未必转化为稳定碳库。此外,有Meta分析研究表明,作物残体移除会显著降低土壤有机碳储量,且该效应受到耕作制度的调节[30]。这意味着“生物量输入—农艺扰动”之间存在显著权衡关系。因此,本研究所观察到的情景差异,本质上反映了土地利用梯度与管理干扰强度的交互作用。
3.3 麻类作物生态价值的不确定性与多维评估
尽管SOC常被作为评价土壤固碳能力的核心指标,但单一化学指标难以全面反映土壤生态过程。Bronick等[31]发现,土壤结构与团聚体形成在有机碳稳定化过程中发挥重要作用,强调物理保护机制对碳保存的重要性,说明碳储量变化与结构变化并非完全同步。同时,微生物生物量碳作为活性碳库的重要组成部分,是碳循环过程的敏感指标。Wardle等[32]指出,MBC反映了土壤生物活性与碳转化速率的变化。因此,仅依赖SOC浓度变化,可能忽视微生物驱动过程与物理结构变化的不同步响应。基于此,本研究采用SOC、BD与MBC三位一体指标体系,从化学储量、物理结构和生物活性三个维度综合评估作物种植对土壤生态系统的影响。这种多维框架有助于避免单指标评价可能带来的过度简化。
3.4 研究局限性与进一步研究策略
本研究的局限性在于现有高质量田间定量研究的数量依然有限,部分数据依赖估算,这在一定程度上增加了不确定性。同时农业土壤碳变化通常具有显著的时间滞后效应,West等[33]在对耕作制度影响SOC的Meta分析中发现,土壤碳库调整往往需要多年甚至更长时间才能稳定显现。因此,短期田间研究可能低估或高估作物种植的真实碳效应。此外,减少耕作扰动被认为是维持碳输入效应的重要管理措施[5]。麻类作物的土壤碳效应受到农艺措施等的综合调控。在高频扰动体系中,即使作物具有较高生物量输入,其碳汇潜力仍可能受到削弱。因此,未来生态工程实践应避免简单地将高生物量作物等同于高固碳潜力,而应结合土地利用背景与管理强度,构建“作物—生境—管理”协同优化策略。在碳亏缺严重的退化或边际土地中,增加植被覆盖和减少扰动可能具有更高的边际收益;而在常规高强度农田中,则需与保护性耕作措施协同实施,以避免农艺干扰抵消潜在的碳输入效应。
4 结论
本研究揭示了麻类作物在农业生态系统中具有较强碳汇能力,但其碳汇效应受到生态梯度及农作制度的影响。高生物量根系形成了麻类作物农田碳源注入的基础,但其土壤碳稳定性受农艺扰动的制约。在干旱、沙漠化等碳亏缺程度较高的边际土地生境下,麻类作物基于高基线差和干预式农田管理措施将产生强修复潜力,而集约化高频扰动将损耗碳输入效益。本研究认为,仅依靠麻类作物高生物量的特性实现土壤固碳不具备全面的可操作性,在固碳生态工程中实施麻类作物干预方案时,应基于生境差异设计“生境-作物-管护”复合型系统技术体系。
参考文献
[1]Quan Q,Zhou J,Hanson P J,et al.Drought-induced peatland carbon loss exacerbated by elevated CO2 and warming[J].Science,2025,390(6771):367-370.
[2]Ohlert T,Smith M D,Collins S L,et al.Drought intensity and duration interact to magnify losses in primary productivity[J].Science,2025,390(6770):284-289.
[3]Lal R.Soil carbon sequestration impacts on global climate change and food security[J].Science,2004,304(5677):1623-1627.
[4]Paustian K,Lehmann J,Ogle S,et al.Climate-smart soils[J].Nature,2016,532(7597):49-57.
[5]Six J,Conant R T,Paul E A,et al.Stabilization mechanisms of soil organic matter:Implications for C-saturation of soils[J].Plant and Soil,2002,241(2):155-176.
[6]Balesdent J,Basile-Doelsch I,Chadoeuf J,et al.Atmosphere-soil carbon transfer as a function of soil depth[J].Nature,2018,559(7715):599-602.
[7]Xu X,Zhang X M,Zou Y Y,et al.Integrated carbon and nitrogen management for cost-effective environmental policies in China[J].Science,2025,388(6751):1098-1103.
[8]Cleophas F N,Zahari N Z,Murugayah P,et al.Phytoremediation:a novel approach of bast fiber plants(hemp,kenaf,jute and flax)for heavy metals decontamination in soil:review[J].Toxics,2023,11(1):5.
[9]Rehman M,Luo D J,Pan J,et al.Sustainable environmental remediation with bast fiber crops:Phytoremediation potential and resource management[J].Science of the Total Environment,2025,977:179403.
[10]Minasny B,Malone B P,McBratney A B,et al.Soil carbon 4 per mille[J].Geoderma,2017,292:59-86.
[11]Page M J,McKenzie J E,Bossuyt P M,et al.The PRISMA 2020 statement: an updated guideline for reporting systematic reviews[J].Bmj,2021:n71.
[12]Wiebe N,Vandermeer B,Platt R W,et al.A systematic review identifies a lack of standardization in methods for handling missing variance data[J].Journal of Clinical Epidemiology,2006,59(4):342-353.
[13]平晓燕,林长存,白宇,等.新疆阿勒泰平原荒漠罗布麻种植区的生态效益评价[J].草业学报,2014,23(2):49-58.
[14]何翠翠,张文,刘立生,等.作物种植对梯田土壤肥力要素演变的影响[J].中国土壤与肥料,2023(5):190-197.
[15]苗菁,宗万松,吕敏,等.基于高通量测序技术研究土壤细菌对盐生植被演替的响应[J].土壤通报,2018,49(3):602-608.
[16]钟义军,武琳,黄欠如,等.不同种植方式对红壤旱地土壤水分及抗旱能力的影响[J].中国水土保持科学,2014,12(2):47-51.
[17]王娟娟,王倩,姜爱霞,等.黄河三角洲盐生植被演替对土壤真菌碳源代谢多样性的影响[J].土壤学报,2018,55(5):1264-1275.
[18]谢瑾,李朝丽,李永梅,等.纳板河流域不同土地利用类型土壤质量评价[J].应用生态学报,2011,22(12):3169-3176.
[19]Liu C,Liu W S,Huot H,et al.Biogeochemical cycles of nutrients,rare earth elements (REEs) and Al in soil-plant system in ion-adsorption REE mine tailings remediated with amendment and ramie (Boehmeria nivea L.)[J].Science of the Total Environment,2022,809:152075.
[20]Zhu H H,Wang S,Zhu Q H,et al.Perennial ramie cropping sustainably increases C sequestration of subtropical upland soils[J].Geoderma,2021,381:114688.
[21]Kumar M,Kundu D K,Ghorai A K,et al.Carbon and nitrogen mineralization kinetics as influenced by diversified cropping systems and residue incorporation in Inceptisols of eastern Indo-Gangetic Plain[J].Soil and Tillage Research,2018,178:108-117.
[22]Kumar M,Mitra S,Mazumdar S P,et al.Improvement of soil health and system productivity through crop diversification and residue incorporation under jute-based different cropping systems[J].Agronomy,2021,11(8):1622.
[23]Kumar M,Mitra S,Mazumdar S P,et al.System productivity,soil carbon and nitrogen sequestration of intensive rice-based cropping systems can be improved through legume crop inclusion with appropriate fertilizer application and crop residues incorporation in the eastern Indo-Gangatic plain[J].Plant and Soil,2025,507(1/2):25-46.
[24]Mohanty S,Saha S,Saha B,et al.Substitution of fertilizer-N with biogas slurry in diversified rice-based cropping systems:Effect on productivity,carbon footprints,nutrients and energy balance[J].Field Crops Research,2024,307:109242.
[25]Parenti A,Zegada-Lizarazu W,Pagani E,et al.Soil organic carbon dynamics in multipurpose cropping systems[J].Industrial Crops and Products,2022,187:115315.
[26]Pude?ko K,Ko?odziej J,Mańkowski J.Restoration of minesoil organic matter by cultivation of fiber hemp (Cannabis sativa L.) on lignite post-mining areas[J].Industrial Crops and Products,2021,171:113921.
[27]Lucas S T,Silvernail A F,Lewis M D.Effects of traditional field retting of hemp on soil organic carbon and the soil microbial community[J].Soil Science Society of America Journal,2022,86(3):742-757.
[28]Poeplau C,Don A.Carbon sequestration in agricultural soils via cultivation of cover crops-A meta-analysis[J].Agriculture,Ecosystems&Environment,2015,200:33-41.
[29]Borenstein M,Hedges L V,Higgins J P T,et al.Introduction to Meta-Analysis[M].Hoboken,NJ:Wiley,2021.
[30]Blanco-Canqui H.Crop residue removal for bioenergy reduces soil carbon pools:how can we offset carbon losses?[J].BioEnergy Research,2013,6(1):358-371.
[31]Bronick C J,Lal R.Soil structure and management:a review[J].Geoderma,2005,124(1/2):3-22.
[32]Wardle D A.A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil[J].Biological Reviews,1992,67(3):321-358.
[33]West T O,Post W M.Soil organic carbon sequestration rates by tillage and crop rotation:a global data analysis[J].Soil Science Society of America Journal,2002,66(6):1930-1946.
文章摘自:兰小明,陶宇峰,颜世伟,高钢,赵信林,朱爱国,陈继康.麻类作物的土壤碳效应:基于Meta与SWiM方法的定量研究[J/OL].中国麻业科学.https://link.cnki.net/urlid/43.1467.S.20260324.1040.008
