摘 要:为评价不同苎麻新品系的镉(Cd)富集转运特性及Cd耐性,研究以中苎1号苎麻品种(Z1)和5个苎麻新品系(H1~H5)为试验材料。通过水培Cd胁迫试验比较分析其植株生长、生物量、Cd富集和转运特性等指标,结合隶属函数法和主成分分析法评价其耐Cd性和Cd富集能力。结果表明:与对照组相比,Cd胁迫显著抑制了苎麻地上部和地下部的生长以及生物量的积累;Z1的叶绿素相对含量(SPAD值)降幅(29.47%)和脯氨酸(Pro)含量升幅(647.54%)最显著。H1的过氧化氢酶(CAT)活性上升最显著(248.86%),Cd转运系数(TF)最高,而Z1的Cd生物富集系数(BCF)最高。综合评价表明,H4的耐Cd性最强,Z1的Cd富集能力最强。试验结果丰富了重金属修复植物可利用资源,为重金属修复专用苎麻新品种选育提供了科学参考。
关键词:苎麻;镉胁迫;生理生化特征;耐镉性评价;镉富集能力评价
镉(Cd)具有很强的生物毒性,容易通过食物链危害人类健康[1]。我国重金属污染严重,根据2014年我国环境保护部和国土资源部发布的《全国土壤污染状况调查公报》显示,全国土壤总的重金属超标率为16.1%,在常见的8种重金属污染物中Cd的超标率最高,达7.0%[2]。因此,环境重金属污染修复是农业安全生产和农业可持续发展的重要课题。植物修复技术凭借其低成本、高效率以及其生态友好等优点从众多修复方法(土壤淋洗、土壤改良剂、化学处理、客土法等)[3]中脱颖而出,越来越受人们的关注。植物修复用的重金属超积累植物有龙葵、东南景天、商陆等[4-6],但这些植物大多为野生型,生长缓慢、生物量小、经济价值低,且只有生长在矿山矿区等地区的生态型植物种群才具有重金属超富集能力和耐受性[7],在实际应用中的效果有限。
苎麻(Boehmeria niveaL.)又叫中国草,是优质天然纤维作物。与Cd超富集植物相比,苎麻具有根系发达、生物量大、抗逆性好、经济价值高等优点,对多种重金属元素有较强的耐性和吸附能力,且对环境中重金属的提取率远高于超积累植物[8],在重金属污染土壤治理中有很大的应用潜力[9]。重金属修复专用苎麻新品种的选育为苎麻治理重金属提供可利用资源,是植物修复重金属领域中重要的环节。前期,以镉耐性和富集能力差异显著的2个苎麻品种作亲本,通过人工杂交选育获得5个优良的新品系(H1、H2、H3、H4和H5)。本研究以耐Cd性和Cd富集能力较强的优质苎麻品种中苎1号(Z1)为对照,采用水培Cd胁迫试验对这5个新品系进行耐Cd性及Cd富集转运特性评价,以期丰富植物修复重金属污染可利用资源种类,为重金属修复专用苎麻新品种选育奠定基础。
1 材料与方法
1.1 试验材料
本研究所用的试验材料为中国农业科学院麻类研究所选育的中苎1号(Z1)苎麻品种,以及5个苎麻新品系(H1、H2、H3、H4和H5)。其中,中苎1号为湖南地区主栽品种,具有产量高、纤维品质好、耐Cd性和Cd富集能力强等优点。5个新品系连续4年大田种植的农艺性状表现稳定、抗逆性好,其中H1和H4的鲜麻和干麻重优于其他3个品系,H2皮厚较高,H3株高最高,H5分蔸能力好,H4纤维细度最高。Cd胁迫试验所用试剂为CdCl2•2.5H2O。
1.2 试验方法
1.2.1 Cd胁迫试验
采集苎麻侧枝,将侧枝末端削尖至15cm左右,仅保留顶端2~3片新叶。将处理好的侧枝放入稀释500倍的多菌灵溶液中浸泡消毒10~20s,随后扦插至预先消毒的水培仪中,每盆15株,每个参试材料2盆。自来水培养至生根,待苎麻水培苗根长至8~10cm时进行Cd处理(Cd2+浓度为40mg/L),以未添加Cd苎麻为对照。每天观察苎麻的生长情况并定期记录数据。试验于室内进行,温度设置为24~26℃,光照强度为300µmol/m2/s,光照周期为14h光照/10h黑暗。试验期间每隔7d向培养液中添加海珀尼卡液肥A液和B液各5mL。
1.2.2. 测定方法
株高测定:处理0、5、10、15d后,测量各植株茎基部到茎顶端的长度。每个材料处理组和对照组各测3个生物学重复,计算株高增长量。
根长测定:处理0、5、10、15d后,测量各植株根系的长度。每个材料处理组和对照组各测3个生物学重复,计算根长增长量。
叶片SPAD值测定:处理7d后使用SPAD-502叶绿素仪测定叶片SPAD值,测定顶端向下第4片完全展开叶,远离叶脉的3个点测量,每个处理每个材料均测定3个生物学重复,取平均值。
生理生化指标测定:处理15d后,采集同一叶位完全展开叶,每盆3个生物学重复。使用茚三酮法测定脯氨酸(Pro)含量,微板法测定丙二醛(MDA)含量,酶联免疫吸附测定法测定金属硫蛋白(MT)含量,氮蓝四唑法测定超氧化物歧化酶(SOD)活性,紫外光比色法测定过氧化物酶(POD)活性以及钼酸铵法测定过氧化氢酶(CAT)活性。所用试剂盒见表1。
表1 试剂盒名称及型号

生物量测定:处理15d后分别收获苎麻地上部和地下部,105℃杀青30min,然后70℃烘至恒重后称量干重。
Cd积累量测定:分别将苎麻地上部和地下部研磨成粉末,过60目筛,采用石墨炉原子吸收光谱法进行测定,参照GB/T17141—1997。
Cd转运系数(TF):地上部Cd积累量/地下部Cd积累量。
Cd生物富集系数(BFC):苎麻单株Cd浓度/Cd处理浓度(40mg/L)。
在试验过程中,Cd处理组各苎麻种质部分植株出现不耐受甚至死亡的现象,导致剩余植株数量不一,为保证植株收获烘干后各苎麻种质样品的干重达到测量Cd含量所需样品重量,将各苎麻种质所剩植株合并,测量单株干重以及Cd富集转运特性指标,不做显著性分析。
1.2.3 数据处理与分析
采用Excel2017处理试验数据,采用IBM SPSS Statistics 27.0.1软件进行相关性分析、方差分析,利用one-way ANOVA进行显著性检验,在p<0.05时认为差异达显著水平,采用Origin Pro 2021进行绘图。通过隶属函数分析和主成分分析对参试材料进行耐Cd性综合评价,指标分类及计算公式如下。
耐Cd性指标:地上部和地下部增长量、地上部和地下部单株干重、SPAD值、Pro含量、MT含量、MDA含量、SOD活性、CAT活性和POD活性,其中Pro和MDA含量采用其值的负数进行分析。
Cd富集能力指标:地上部和地下部Cd浓度、地上部和地下部单株总Cd积累量、Cd转运系数和Cd生物富集系数。
对耐Cd性指标和Cd富集能力指标中各个单项指标进行主成分分析,提取累计贡献率85%以上的综合指标(i个),随后进行隶属函数分析。
U(Xi)=(Xi―Ximin)/(Ximax―Ximin)
式中:U为隶属函数值;Xi为第i个综合指标;Ximin为所有综合指标中的最小值;Ximax为所有综合指标中的最大值;i=1,2,3,…,n。

式中:Wi为第i个综合指标在所有综合指标中的权重;Pi为第i个综合指标的贡献率。

式中:D值为Cd胁迫下各参试材料的耐Cd性和Cd富集能力综合评价值;i=1,2,3,…,n。
2 结果与分析
2.1 Cd胁迫对不同苎麻新品系生长的影响
从图1中可看出,对照组中(未添加Cd),处理5d后5个新品系的株高增长量均低于对照品种Z1,而在处理10d后H2和H3地上部和地下部的生长明显优于Z1,表现为其株高的增长量分别比Z1高32.08%和41.51%,根长的增长量分别比Z1高29.05%和14.68%。除H1外,H2~H5的生物量积累均高于Z1,其中H3的生物量最高,其次是H2,分别比Z1高50.28%和40.11%。在Cd胁迫后,各苎麻种质植株均出现失水萎缩的现象,部分植株甚至出现枯萎死亡的现象,导致株高和根长增长量出现负增长。
与对照组相比,Cd胁迫显著抑制了6个参试材料地上部和地下部的生长,各参试材料植株地上部均出现失水萎蔫的现象,生物量积累降低。当处理时间低于10d时,H1~H4的株高增长量低于对照品种Z1,而Cd胁迫15d后,其株高增长量高于Z1。与其他4个新品系相比,H5地上部受到Cd毒害最小,萎蔫程度最小。Cd胁迫下,5个新品系的根长增长量均高于Z1,其中H1的增长量最高,比Z1高203.43%。6个参试材料在Cd处理15d后的生物量积累从高到低排序为H1>H4>H5>Z1>H3>H2。

注:不同小写字母表示相同处理时间同一Cd浓度下各参试材料与Z1相比的数值差异显著(p<0.05)。Z1表示中苎1号,H1—H5表示5个新品系。下同
图1 Cd胁迫对苎麻生长的影响
2.2 Cd胁迫下不同苎麻新品系叶片SPAD值变化特征
从图2中可看出,对照组中H1、H2、H4和H5的SPAD值略低于对照品种Z1,H3的SPAD值显著低于Z1。Cd胁迫5d后,Z1、H2和H3的大部分植株叶片(分别为8株、10株和10株)开始出现失水萎蔫现象,在处理中期(7~10d)各参试材料植株叶片均呈现发黄甚至脱落的症状。参试的6个材料SPAD值与对照组相比呈下降趋势,其中Z1的SPAD值下降幅度最大(下降了29.47%),5个新品系下降的幅度小,其SPAD值从高到低排序为H1>H5>H4>H2>H3>Z1。
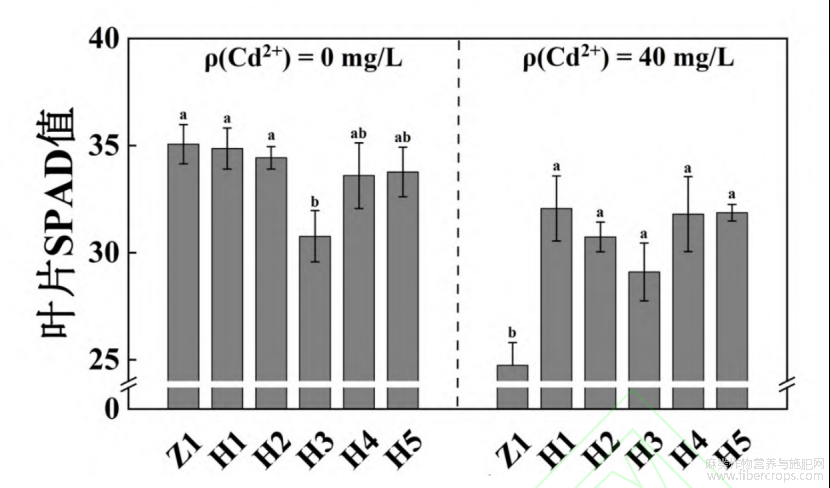
图2 Cd胁迫对苎麻叶片SPAD值的影响
2.3 Cd胁迫下不同苎麻新品系叶片抗氧化体系组分差异
对各个参试材料的叶片抗氧化体系组分进行了分析,结果发现,在不加Cd的条件下H1的Pro含量比对照品种Z1高67.6%,而H2、H3和H5的Pro含量显著低于Z1(图3-A)。H2和H3的MT含量显著高于对照品种Z1,而H1、H4和H5的MT含量显著低于Z1(图3-B)。H1和H2的MDA含量略高于对照品种Z1,而H3、H4和H5分别比Z1低23.53%、11.16%和12.88%(图3-C)。H1和H2的SOD活性显著高于对照品种Z1和H3、H4、H5,H3的SOD活性显著低于Z1(图3-D)。H5的CAT活性显著高于Z1和其他4个品系,而H1和H2的CAT活性显著低于Z1(图3-E)。除H4以外的4个新品系的POD活性比Z1高1.48%~24.45%,H1、H2和H5的POD活性与Z1相比达显著水平(图3-F)。
Cd胁迫下,与对照组相比,Z1、H2、H3和H5的Pro含量和Z1、H1、H2、H4和H5的MT含量显著升高,其中Z1的Pro含量升高幅度最显著,达647.54%,5个新品系的Pro含量均显著低于对照品种Z1(图3-A和图3-B)。Cd胁迫还增加了6个参试材料的MDA含量以及SOD活性(比对照组提升了45.46%~78.63%)和POD活性(比对照组提升了7.37%~45.49%),其中H1和H2的MDA含量和SOD活性显著高于Z1(图3-C和图3-D),而H3的SOD活性显著低于Z1,H1和H4的POD活性显著高于Z1(图3-F)。受Cd胁迫诱导,H1的CAT活性与其对照组相比提升了248.86%,且显著高于对照品种Z1,而其他4个品系的CAT活性在Cd胁迫下显著低于Z1,其中H2的CAT活性最低,其次是H5(图3-E)。

图3 Cd胁迫对苎麻叶片抗氧化体系组分的影响
2.4 不同苎麻新品系Cd富集转运特性
从图4中可看出,H1~H5新品系的地下部和地上部Cd浓度均低于对照品种Z1,其地上部和地下部Cd浓度分别比Z1低5.68%~74.56%和4.46%~97.05%(图4-A)。5个新品系的单株总Cd富集量也均低于Z1,其地上部(H3地上部Cd积累量略高于Z1)和地下部Cd积累量分别比Z1低7.30%~45.37%和20.07%~65.59%(图4-B)。5个新品系的Cd浓度和单株Cd富集量由高到低顺序为H3>H4>H2>H1>H5。
H1、H2、H3、H5的转运系数(TF)分别比对照品种Z1高132.86%、132.52%、65.80%和17.56%,而H4的TF比Z1低19.23%,其中H1和H2的TF最高,分别是1.305和1.303(图4-C)。5个新品系对Cd的生物富集系数(BCF)均小于Z1,BCF从高到低的顺序为Z1>H3>H2>H4>H5>H1(图4-D)。
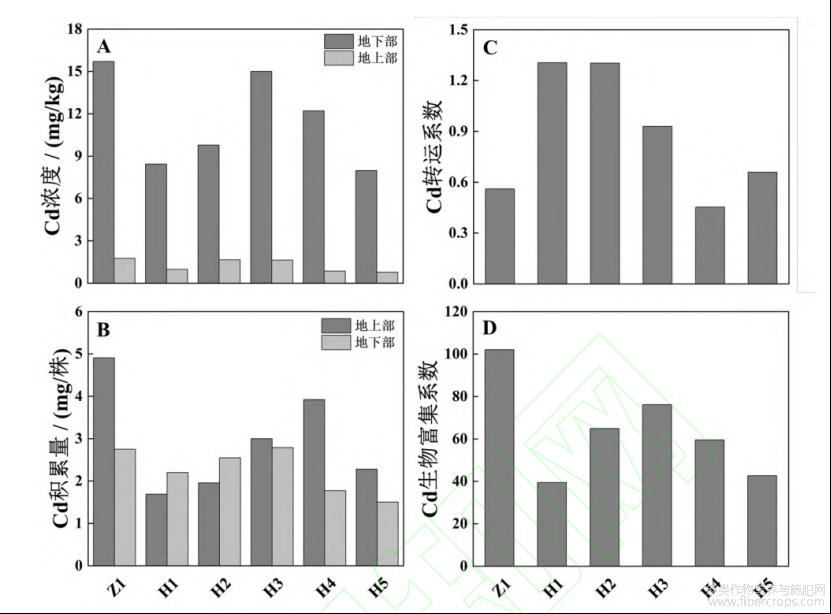
图4 Cd胁迫对苎麻Cd富集转运特性的影响
2.5 各指标相关性分析
利用SPSS对多个指标进行相关性分析,结果如图5所示,在Cd胁迫下各指标之间有着不同程度的相关性。其中地下部增长量与POD活性呈显著正相关,说明在Cd胁迫下Cd耐性强的苎麻可通过响应POD保护机制,缓解Cd对苎麻地下部的毒害作用;Cd转运系数与叶片中的MDA含量呈极显著正相关,说明在Cd胁迫下,Cd转运能力强的苎麻会将根系吸附的Cd离子转运至地上部,导致植株叶片中MDA含量升高,造成膜质过氧化损伤;此外Cd富集系数与叶片SPAD值呈极显著负相关,但与Pro含量呈显著正相关,其中叶片SPAD值与Pro含量呈显著负相关。
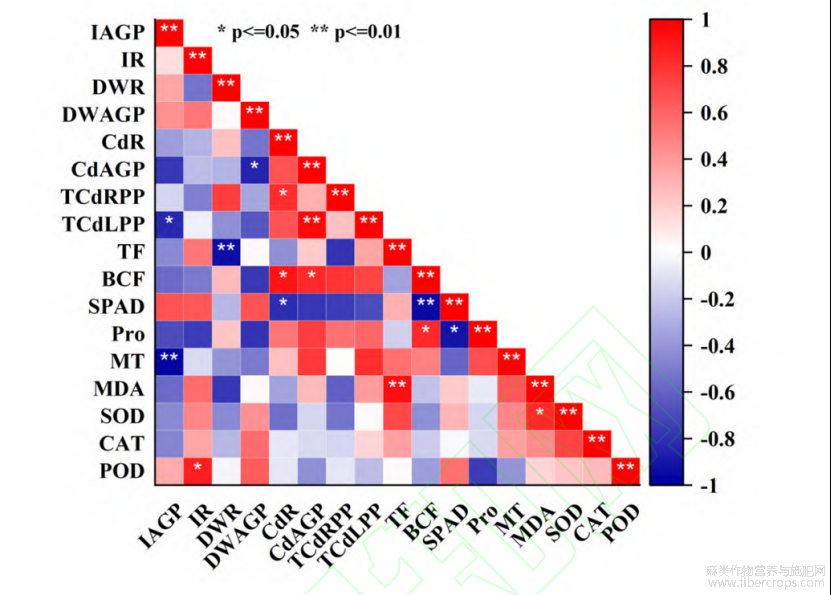
注:IAGP—地上部增长量;IR—地下部增长量;DWR—地下部单株干重;DWAGP—地上部单株干重;CdR—地下部单株Cd浓;CdAGP—地上部单株Cd浓度;TCdRPP—地下部单株总Cd积累量;TCdLPP—地上部单株总Cd积累量;TF—Cd转运系数;BCF—Cd生物富集系数;SPAD—叶片SPAD值;Pro—脯氨酸含量;MT—金属硫蛋白含量;MDA—丙二醛含量;SOD—超氧化物歧化酶活性;CAT—过氧化氢酶活性;POD—过氧化物酶活性。
图5 Cd胁迫下苎麻各项指标皮尔逊相关系数热图
2.6 不同参试材料耐Cd性综合评价
通过主成分分析从耐Cd性指标组中提取出累计贡献率85%以上的3个新的综合指标,分别是PC1、PC2和PC3,其贡献率分别为43.25%、35.88%和11.31%(表2)。综合评价D值为各参试材料的耐Cd能力,D值越大表示该苎麻生长受Cd毒害的影响越小。结果显示,耐Cd性由强到弱依次为H4>H5>H1>H3>H2>Z1(表3)。
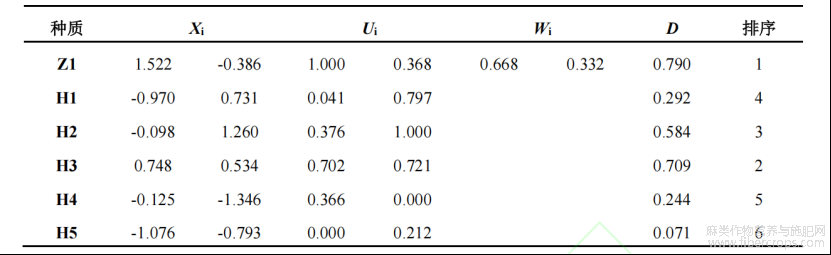
Cd富集能力指标组中提取出累计贡献率85%以上的两个新的综合指标,分别是PC1和PC2,其贡献率分别为64.63%和32.17%(表4)。其综合评价D值为各参试材料的Cd富集能力,D值越大表示该苎麻的Cd富集能力越强。结果显示,Cd富集能力由强到弱依次为Z1>H3>H2>H1>H4>H5(表5)。
表2 耐Cd性各综合指标的系数及贡献率
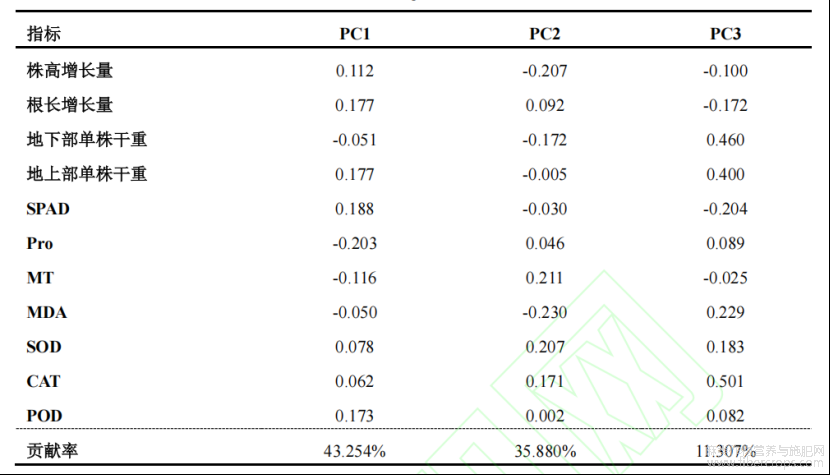
表3 不同参试材料综合指标值、隶属函数值、综合指标权重、综合评价值和耐Cd性综合排序
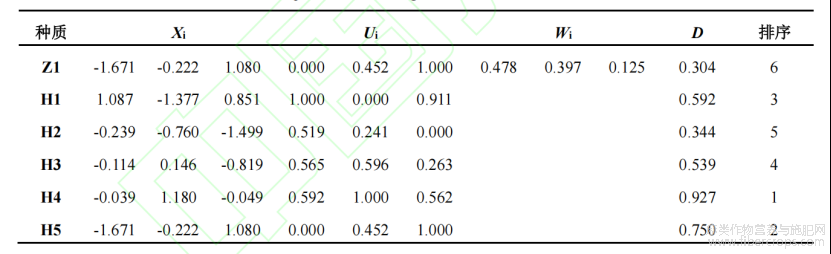
表4 Cd富集能力各综合指标的系数及贡献率
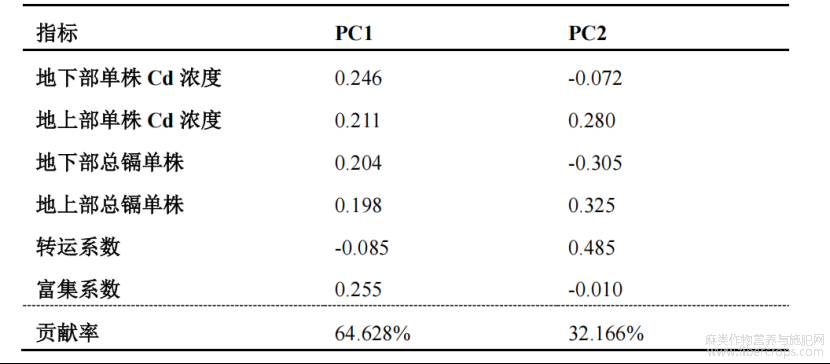
表5 不同参试材料综合指标值、隶属函数值、综合指标权重、综合评价值和Cd富集能力综合排序
3 讨论
植物体内Pro含量的增加是植物应对逆境胁迫所产生的一种生理生化反应,其通过渗透调节[10]、调节细胞氧化还原电位[11]以及清除活性氧[12]等方式提高植物抗逆性。另有研究表明,Pro是反映植物细胞结构和功能受到伤害[13-14]的指标。在本试验中:Cd胁迫下Z1、H2、H3和H5苎麻体内的Pro含量与对照组相比显著升高,其中Z1苎麻最为显著;而Cd胁迫下的SPAD值显著低于对照组,且Z1苎麻下降的幅度最大。据此推测,Cd胁迫下Pro含量增加表明苎麻细胞结构和功能受损,从而导致叶绿素的合成受到抑制。SPAD值与Pro含量之间的显著负相关进一步证实了这一推测。并且,Z1苎麻的根长增长量和生物量积累是6个参试材料中最低的,结合以上结果可看出,Cd胁迫下植株叶片中Pro含量高的苎麻从侧面反映出其根系生长和生物量积累受到抑制。因此在本试验条件下,Pro含量可作为反映苎麻植株在Cd胁迫下受到伤害的指标,在进行耐Cd性综合评价时需将Pro含量数据转化成相反数再进行主成分分析和隶属函数分析。
SOD是重要的抗氧化酶之一,SOD可催化O2−生成H2O2和O2,从而清除植物体内的自由基,是重要的活性氧防御酶[15]。但其在清除自由基的同时也会产生H2O2,而H2O2的积累会作为一种胁迫因子破坏细胞完整性,降低保护酶的活性,从而影响细胞正常的生理活动[16]。在植物中负责清除H2O2的主要有CAT和POD两种抗氧化酶[17],CAT主要负责将植物体内高浓度的H2O2清除,使其浓度维持在一个较低的水平[18],而POD则是把低浓度的H2O2转化为H2O,两者相结合起到清除H2O2保护植物的目的。前人研究结果表明[19],Cd浓度是影响植物体内抗氧化酶活性的重要因素之一,具体表现为低浓度Cd胁迫促进抗氧化酶活性,高浓度Cd胁迫抑制抗氧化酶活性[20]。在本试验中,与对照组相比,Cd胁迫下6个参试材料的SOD和POD活性均有不同程度的增加,其中H1苎麻的SOD活性和H4苎麻的POD活性增加幅度最高。而Cd胁迫后除H1苎麻外其他参试材料植株叶片中的CAT活性均受到抑制,其中H5苎麻的CAT活性下降最为显著,而H1苎麻的CAT活性则是大幅增加。这说明在相同条件下植物的种质也是影响抗氧化酶活性的重要原因之一。
当植物处于高温、干旱、重金属等胁迫环境下时,植物的生长会受到不同程度的损害,通过调节体内的生理生化过程以适应逆境,形成一定的抗逆机制。而苎麻作为多年生宿根植物,生长迅速、生物量大,并且较多研究表明,苎麻具有较强的Cd富集能力和耐受性,但不同品种间差异较大[9]。此外有研究[21]表明,植物的生物量与植物对Cd的积累量呈显著负相关,Cd的转运系数主要用于评价植物将Cd从根部运输到地上部的能力,而Cd在根中的固定有助于提高植物的耐Cd性[22],意味着耐Cd性强的品种其Cd转运系数一般都低[23]。在本试验的相关性分析中,Cd富集系数与SPAD值呈极显著负相关,与Pro含量呈显著正相关,说明Cd富集能力强的苎麻由于体内富集了较多的Cd导致其叶绿素合成受到显著抑制,细胞结构和功能受到破坏;Cd转运系数与根干重呈极显著负相关,与MDA含量呈极显著正相关,说明Cd转运能力强的苎麻其根部生物量显著低于其他苎麻,且叶细胞膜质过氧化程度高。结合耐Cd性和Cd富集能力综合评价D值来看,耐Cd性最强的H4和H5苎麻,其Cd富集能力是最差的,这与前人的研究结果[21-23]类似。本试验通过隶属函数分析和主成分分析法将17个指标降维,并消除了各指标间复杂的相关性,提高了模型的准确性,最后得出综合评价D值,使这6个参试材料的耐Cd性和Cd富集能力评价更为直观和科学。D值的计算结果表明:在水培环境40mg/L的Cd2+条件下,H1苎麻的耐Cd性最强,Z1最弱;Z1苎麻的Cd富集能力最强,H5苎麻最弱。前人研究[24]表明,Z1苎麻具有较强的耐Cd性,而本试验中的H1~H5苎麻虽然Cd富集能力比Z1弱,但耐Cd性都比Z1强,这也从侧面说明了这5个苎麻新品系在重金属修复领域有着很好的应用潜力,但其大田重金属修复能力还需要进一步的研究和验证。
4 结论
综上所述,6个参试材料的耐Cd性由强到弱依次为H4>H5>H1>H3>H2>Z1,而Cd富集能力由强到弱依次为Z1>H3>H2>H1>H4>H5。本试验结果丰富了重金属修复植物可利用资源种类,为重金属修复专用苎麻新品种选育提供科学参考。
参考文献
[1]LUO H F,ZHANG J Y,JIA W J,et al.Analyzing the role of soil and rice cadmium pollution on human renal dysfunction by correlation and path analysis[J].Environmental Science and Pollution Research,2017,24(2):2047-2054.
[2]陈能扬,郑煜基,何晓峰,等.《全国土壤污染状况调查公报》[J].中国环保产业,2014(5):2.
[3]KHALID S, SHAHID M,NIAZI N K,et al.A comparison of technologies for remediation of heavy metal contaminated soils[J].Journal of Geochemical Exploration,2017,182:247-268.
[4]魏树和,周启星,王新,等.一种新发现的镉超积累植物龙葵(Solanum nigrum L)[J].科学通报,2004,49(24):2568-2573.
[5]杨肖娥,龙新亮,倪吾钟,等.东南景天(Sedum alfredii H):一种新的锌超积累植物[J].科学通报,2002,47(13):1003-1006.
[6]薛生国,陈英旭,林琦,等.中国首次发现的锰超积累植物:商陆[J].生态学报,2003,23(5):935-937.
[7]杨肖娥,龙新亮,倪吾钟,等.古老铅锌矿山生态型东南景天对锌耐性及超积累特性的研究[J].植物生态学报,2001,25(6):665-672.
[8]杨勇,王巍,江荣风,等.超累积植物与高生物量植物提取镉效率的比较[J].生态学报,2009,29(5):2732-2737.
[9]曹德菊,周世怀,项剑.兰麻对土壤中镉的耐受和积累效应研究[J].中国林业,2004,26(6):272-274.
[10]CHRISTIAN J.The influence of nutrition on the water relations of Salmonella oranienburg[J].Australian Journal of Biological Sciences,1955,8(1):75.
[11]SARADHI P P.Proline accumulation under heavy metal stress[J].Journal of Plant Physiology,1991,138(5):554-558.
[12]SMIRNOFF N. The role of active oxygen in the response of plants to water deficit and desiccation[J].New Phytologist,1993,125(1):27-58.
[13]汤春城.逆境条件下植物脯氨酸的累积及其可能的意义[J].植物生理学通讯,1984,20(1):15-21.
[14]卞咏梅,陈树元,刘绍考,等.大气氟污染对某些植物脯氨酸的影响[J].植物生理学通讯,1988,24(6):19-21.
[15]黄玉山,罗广华,关宗文.镉诱导植物的自由基过氧化损伤[J].植物学报(英文版),1997,39(6):522-526.
[16]林植芳,李双顺,林桂珠,等.衰老叶片和叶绿体中H?O?的累积与膜脂过氧化的关系[J].植物生理学报,1988,14(1):16-22.
[17]杨淑娟,高俊凤,汪佳军.自由基与植物的衰老[J].西北植物学报,2001,21(2):215-220.
[18]潘瑞炽,豆志杰,叶庆生.茉莉酸甲酯对水分胁迫下花生幼苗SOD活性和膜脂过氧化作用的影响[J].植物生理学报,1995,21(3):221-228.
[19]李洋,于丽杰,金晓霞.植物重金属胁迫耐受机制[J].中国生物工程杂志,2015,35(9):94-104.
[20]吴嘉煜,米楠.重金属对植物抗氧化酶影响研究进展[J].浙江农业科学,2022,63(6):1177-1181.
[21]ZHANG X F,GAO B,XIA H P.Effect of cadmium on growth,photosynthesis,mineral nutrition and metal accumulation of Bana grass and vetiver grass[J].Ecotoxicology and Environmental Safety,2014,106:102-108.
[22]CHEN G C,LIU Y Q,WANG R M,et al.Cadmium adsorption by willow root:the role of cell walls and their subfractions[J].Environmental Science and Pollution Research,2013,20(8):5665-5672.
[23]FAN W R,YU H,YAN J Y,et al.Variety-dependent responses of common tobacco with differential cadmium resistance:Cadmium uptake and distribution,antioxidative activity,and gene expression[J].Ecotoxicology and Environmental Safety,2024,281:116596.
[24]牟攀,陈坤梅,朱爱国,等.镉胁迫下苎麻水培苗生理生化变化特征[J].中国麻业科学,2021,43(6):294-302.
文章摘自:赵亿嘉,邱孝军,朱爱国,陈平,陈继康,高钢,王晓飞,陈坤梅.不同苎麻新品系镉富集转运特性及镉耐性评价[J/OL].中国麻业科学.https://link.cnki.net/urlid/43.1467.S.20250928.1021.002
