摘 要:夏季高温环境易影响公牛精液品质及肠道微生物组成和丰度。为探究饲粮添加亚麻籽对热应激公牛生理生化、精液品质及后肠道菌群的影响,为改善热应激公牛精液品质、探究肠道微生物与公牛生育能力的潜在关系提供参考依据,本试验选择年龄3~5岁健康状况良好的西门塔尔种公牛20头,分为对照组和亚麻籽组,每组10头,对照组饲喂基础饲粮,亚麻籽组饲喂基础日粮+亚麻籽120g/d,预试期7d,正试期60d。试验过程中采集牛舍温湿度数据、牛的呼吸频率和直肠温度;测定公牛精子活力、密度、质膜完整率和畸形率,最后一天采集公牛血液和直肠粪便样品送检。结果显示:(1)亚麻籽组公牛的呼吸频率显著降低(P<0.05),而直肠温度在对照组与亚麻籽组之间无显著差异(P>0.05);(2)与对照组相比,亚麻籽组公牛精子畸形率显著降低(P<0.05),质膜完整率显著增加(P<0.05),但精子密度和精子活力无显著变化(P>0.05);(3)公牛血清中丙二醛(malondialdehyde,MDA)、皮质醇(corti-sol,COR)水平显著降低(P<0.05),过氧化氢酶(catalase,CAT)、总抗氧化能力(total an-tioxidantcapacity,T-AOC)水平显著增加(P<0.05);(4)在门水平上,与对照组相比,亚麻籽组中厚壁菌门显著减少(P<0.05),拟杆菌门和螺旋体门显著增加(P<0.05);(5)在属水平,对照组具有显著作用的菌有厌氧菌科(Anaerovoracaceae)、短优杆菌属(Eubacterium_brachy_group),亚麻籽组具有显著作用的菌主要是理研菌科(Rikenellaceae)和巴恩斯氏菌科(Bar-nesiellaceae)。可见,亚麻籽具有抗氧化作用,可减少氧化应激对精子的影响,提高精液品质,且亚麻籽可能通过调节肠道微生物区系进而提高机体对高温环境的耐受性从而调控精液品质。
关键词:亚麻籽;热应激;精液品质;粪便微生物菌群;西门塔尔种公牛
热应激严重影响公牛的生殖性能,高温致使睾丸产生过量的活性氧(reactive oxygen species,ROS),进而与精子质膜中不饱和脂肪酸发生脂质过氧化反应,细胞内线粒体内膜、精子质膜完整性、精子头部及尾部被破坏,造成精液品质下降,影响受精效果[1]。亚麻籽含有大量的亚油酸和α-亚麻酸,是天然的抗氧化剂,其中包含的omega-3和omega-6可保护睾丸质膜免受脂质过氧化(lipid peroxida-tion,LPO)和活性氧的侵害,缓解热应激造成的精液品质下降[2]。热应激公牛饲粮添加亚麻籽可减少脂质过氧化,增加细胞活力、膜流动性和精子顶体完整性[2]。在公牛精子中,精子膜的脂质双分子层主要由磷脂(phospholipids,PLs)和胆固醇组成,细胞膜磷脂含有大量长链多不饱和脂肪酸(long-chain polyunsaturated fatty acids,LC-PUFAs),特别是n-6系列,含有二十二碳五烯酸。LC-PUFAs和胆固醇一起调控精子膜流动性、顶体反应和精卵结合[3]。精子膜上的LC-PUFAs主要来源于必需亚油酸和α-亚麻酸,而亚麻籽含有多种不饱和脂肪酸,如α-亚麻酸[4]。由于反刍动物缺乏相关的脂肪酸去饱和酶,不能合成omega-3 PUFAs,因而反刍动物必须从食物中获取omega-3 PUFAs或其前体[5]。饲粮中添加亚麻籽可以提高动物组织中α-亚麻酸和omega-3的浓度,这2种物质参与重要生殖激素的合成,如17β-雌二醇、促卵泡素和促黄体素等[6-8]。此外,omega-3 PUFAs还含有硬脂酸、油酸、亚油酸和棕榈酸以及酚类化合物,这些化合物在精子发生和抗氧化酶活性中都有重要作用[5]。因此,亚麻籽具有较高的潜在抗氧化作用,保护和刺激抗氧化酶的活性[9]。
热应激增加了呼吸速率和皮肤温度,降低了瘤胃挥发性脂肪酸的分解速率,肠道菌群可能通过改变代谢物或改变消化和能量吸收来改变宿主代谢,从而干扰配子的形成[10]。雄性动物的精子活力与宿主代谢组、睾丸功能和肠道微生物调节之间存在密切联系[11]。Wang等[12]研究指出,不同精子存活率的绵羊肠道微生物和代谢物存在显著差异。肠道微生物群通过调节类固醇性激素、胰岛素敏感性、免疫系统和性腺微生物群的循环水平来促进性腺功能,已有研究证明肠道微生物-性腺轴的存在,以及该轴在性腺功能中的作用[13]。随着16SrRNA基因测序的应用,热应激下公牛生理生化、精液品质和微生物组之间的潜在相关性受到广泛关注。因此,本研究对公牛生理生化、精液品质及粪便微生物进行分析,以期初步揭示亚麻籽与公牛生育能力及肠道健康之间的潜在关系。
1 材料与方法
1.1 试验时间和地点
试验于2023年7月10日至9月18日在河南省鼎元种牛育种有限公司进行,整个试验期采用相同的管理制度,所有饲养动物单圈饲养。
1.2 试验设计及试验日粮
选取年龄3~5岁、健康状况良好的西门塔尔种公牛20头,体重约为1116.38±103.03kg,随机分为2组,每组10头。预试期7d,正试期60d。对照组饲喂基础饲粮,饲粮参照NRC(2001)[14]配制,其饲粮组成及营养水平见表1;亚麻籽组饲喂基础饲粮+120g/d亚麻籽。所有牛每日精饲料饲喂2次(5:30、14:30),粗料饲喂3次(5:30、14:30、19:30)。
14:30饲喂时把亚麻籽添加在混合精料里对试验组的公牛进行单独饲喂。
表1 基础饲粮组成及营养水平(风干基础)
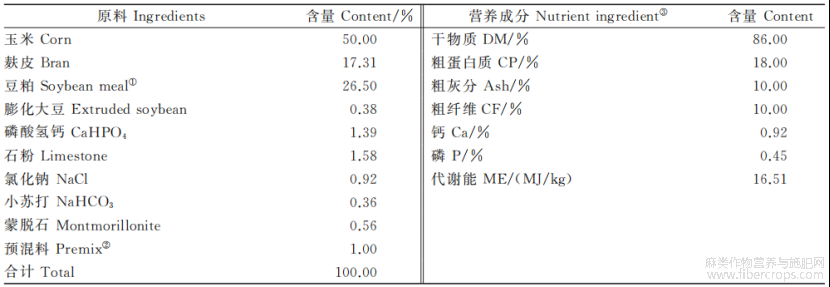
注:①豆粕蛋白主要含量为44.2%,由河南中荷云星饲料有限公司提供。②每千克预混料含:锰800~1000mg,铜200~500mg,铁800~1000mg,锌1000~2000mg,维生素A30000~50000IU,维生素D90000~120000IU。③代谢能为计算值,其他均为实测值。
1.3 牛舍温湿度的测定与统计
试验过程中距离地面垂直高度1.8m处安装温湿度仪,每隔2h自动测定和记录牛舍温度(℃)和相对湿度(%)数据,并转换为温湿度指数(tempera-ture and humidity index,THI)。采用Ingraham的公式计算THI值:THI(%)=(1.8×T+32)-(0.55-0.55×RH)×(1.8×T-26),其中T为摄氏度(℃),RH为相对湿度。
1.4 直肠温度和呼吸频率的测定
在试验期间,每天12:00-14:00,通过目测胸腹部的起伏动作测定每头种公牛1min的腹部起伏次数,连续测3次取其平均值,即呼吸频率。于14:30-15:30,使用兽用电子体温计插入种公牛肛门内,1min后读取温度,即直肠温度。
1.5 血清抗氧化指标和激素水平测定
在试验结束当天每组选择种公牛各6头,并于中午饲喂前每头牛尾根静脉采血10mL,静置30min后进行离心(3500r/min,15min),吸取血清分装于0.5mL离心管中,-20℃保存待测。超氧化物歧化酶(superoxide dismutase,SOD)、MDA、T-AOC和CAT均采用南京建成生物公司试剂盒进行测定;其他血清生化指标,HSP 70使用ELISA试剂盒进行定量检测,使用酶标分析仪(上海酶联生物科技有限公司,型号:ML-dr 3518)测定吸光度(OD值),通过标准曲线计算样品中HSP70的浓度;COR的检测试剂盒使用上海酶联生物科技有限公司生产,其余检测方法同上。
1.6 精液品质测定
利用假阴道法采集精液,并迅速送入实验室进行精液品质检测,每周采精2~3次,记录采精公牛的牛号、射精量、精子活力和精子密度,根据《牛冷冻精液》(GB 4143-2022)[15]的方法测定精子顶体完整率和畸形率,采用精子尾部低渗肿胀法(hypoton-ic swelling of sperm tai,HOST)[16]检测精子质膜完整率。
精子畸形率(%)=(畸形精子数/精子总数)x100%
精子顶体完整率(%)=顶体完整的精子/精子总数x100%。
精子质膜完整率=质膜完整的精子/计数的总精子数×100%。
1.7 直肠粪便采集
在试验的最后一天从公牛直肠采集粪便,放入无菌离心管中液氮速冻,-80℃保存,用于后续16SrRNA基因测序分析。
1.8 DNA抽提和PCR扩增
按E.Z.N.A.![]() soilDNA kit说明书抽提微生物群落总DNA,1%琼脂糖凝胶电泳检测质量,Nano-Drop2000测浓度与纯度。使用338F(5'-ACTC-CTACGGGAGGCAGCAG-3')和806R(5'-GGAC-TACHVGGGTWTCTAAT-3')对16SrRNA基因V3~V4可变区进行PCR扩增,扩增程序如下:95℃预变性3min,27个循环(95℃变性30s,55℃退火30s,72℃延伸30s),72℃延伸10min,最后在4℃进行保存(PCR仪为ABIGeneAmp
soilDNA kit说明书抽提微生物群落总DNA,1%琼脂糖凝胶电泳检测质量,Nano-Drop2000测浓度与纯度。使用338F(5'-ACTC-CTACGGGAGGCAGCAG-3')和806R(5'-GGAC-TACHVGGGTWTCTAAT-3')对16SrRNA基因V3~V4可变区进行PCR扩增,扩增程序如下:95℃预变性3min,27个循环(95℃变性30s,55℃退火30s,72℃延伸30s),72℃延伸10min,最后在4℃进行保存(PCR仪为ABIGeneAmp![]() 9700型)。PCR反应体系为:5×TransStart FastPfu缓冲液4μL,2.5mmol/L dNTPs2μL,上游引物(5μmol/L)
9700型)。PCR反应体系为:5×TransStart FastPfu缓冲液4μL,2.5mmol/L dNTPs2μL,上游引物(5μmol/L)
0.8μL,下游引物(5μmol/L)0.8μL,TransStart Fast-Pfu DNA聚合酶0.4μL,模板DNA10ng,ddH2O补足20μL。每个样本3个重复。
1.9 IlluminaMiseq测序
将同一样本PCR产物混合后,经2%琼脂糖凝胶回收并纯化(AxyPrep Kit),再电泳检测。纯化产物用Quantus Fluorometer定量。使用NEXTflex RapidDNA-SeqKit建库:接头连接、磁珠筛选去自连、PCR扩增富集模板、磁珠回收得文库。文库在IlluminaMiseqPE300上测序(上海美吉)。
1.10 16SrRNA基因测序数据处理
使用FASTP进行质控,去除reads尾部质量值小于20的碱基,窗口50bp内平均质量值小于20的则截断,过滤短于50bp及含N碱基的reads。用FLASH软件根据至少10bp overlap拼接PE reads,允许overlap区20%的错配。通过barcode(0错配)和引物(≤2错配)区分调整序列。最后使用UPARSE以97%相似度聚类OTU并去嵌合体。RDP classifier以70%阈值比对Silva 16S rRNA数据库进行物种注释。
1.11 数据处理分析
数据使用Excel 2021进行初步整理,使用SPSS23.0软件中one-way ANOVA单因素方差分析,Duncan氏进行多重比较。数据以平均值±标准差表示,P<0.05表示差异显著。
2 结果与分析
2.1 牛舍温湿度
基于奶牛热应激评价标准,当THI>72时,牛处于热应激状态。图1所示,在试验期前30d牛舍THI变化范围在78~85,试验期后30d,THI变化范围在70~85,本试验THI平均值78以上,说明试验期间牛均处于热应激状态,且大部分时间处于中度热应激范围。
2.2 亚麻籽对热应激西门塔尔牛直肠温度和呼吸频率的影响
由表2可知,与对照组相比,热应激环境下亚麻籽组牛的呼吸频率显著降低(P<0.05),但是添加亚麻籽对直肠温度无明显改善作用。
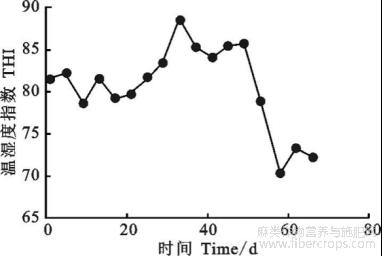
图1 牛舍温湿度指数变化曲线
表2 亚麻籽对热应激西门塔尔牛直肠温度和呼吸频率的影响
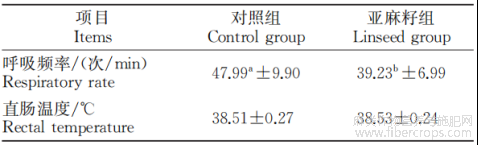
注:同行数据肩标为相同字母或无字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05)。下同。
2.3 亚麻籽对热应激西门塔尔牛血清抗氧化及激素指标的影响
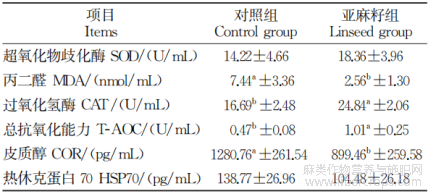
由表3可知,与对照组相比,日粮中添加亚麻籽能够显著提升血清T-AOC、CAT的水平和降低MDA、COR含量(P<0.05)。HSP70、COR是应激损伤程度的生物标志物,对照组HSP70、COR高说明热应激损伤更严重,亚麻籽组HSP70、COR降低则表明其减轻了热应激损伤。表明亚麻籽组可减少应激损伤、改善机体状态、提升抗应激能力。
表3 亚麻籽对热应激西门塔尔牛血清抗氧化及激素的影响
2.4 亚麻籽对热应激西门塔尔牛精液品质的影响
由表4可知,饲料添加亚麻籽对射精量、精子密度、精子活力、总精子数和顶体完整率无显著影响(P>0.05)。与对照组相比,添加亚麻籽种公牛的精子畸形率显著降低、质膜完整率显著增加(P<0.05)。
2.5 16SrRNA基因分析后肠道菌群
2.5.1 α多样性分析 使用Shannan、Simpson、Ace、Chao、Coverage和Sobs指数评估粪便微生物群落的丰富度和多样性。由表5可知,饲喂亚麻籽后,与对照组相比,亚麻籽组的Shannan、Simpson、Ace、Chao、Coverage和Sobs指数无明显差异(P>0.05)。
表4 亚麻籽对热应激西门塔尔牛精液品质的影响
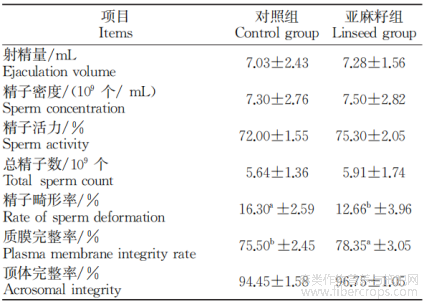
表5 亚麻籽对热应激西门塔尔牛粪便微生物alpha多样性指数的影响
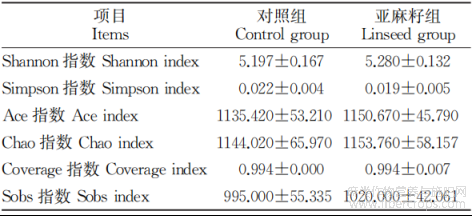
2.5.2 β多样性分析 采用主坐标分析(PCoA)探究亚麻籽对热应激公牛粪便微生物β多样性的影响。如图2所示,在OTU水平上,通过PER-MANOVA检验,表明对照组和亚麻籽组属水平菌群组成差异不显著(P>0.05),对照组与亚麻籽组样本未明显区分。
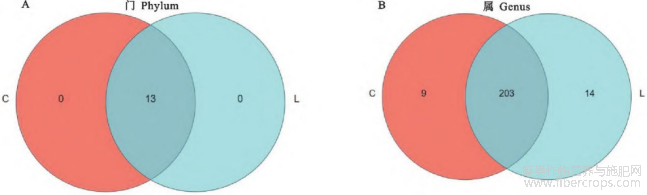
2.5.3 粪便微生物组成及差异分析 对照组和亚麻籽组共有13个门,203个属,对照组特有9个属;亚麻籽组特有14个属(图3);分析对照组和亚麻籽组在门和属水平上丰度前十的微生物物种。在门水平上,厚壁菌门(Firmicutes)、拟杆菌门(Bacteroid ota)和螺旋菌门(Spirochaetota)的丰度较高,是对照组和亚麻籽组粪便微生物群落中共有的优势菌群(图4)。在属水平上,优势菌属有瘤胃球菌科UCG-005属、Rikenellaceae_RC9_gut_group、norank_f_UCG-010、克里斯滕森氏菌(Christensenellaceae_R-7_group)、norank_f_Eubacterium_coprostan-oligenes_group、另枝杆菌属(Alistipes)、密螺旋体属(Treponema)、拟杆菌属(Bacteroides)等(图4)。
由图5可见:在门水平上,与对照组相比,亚麻籽组中厚壁菌门显著减少,拟杆菌门和螺旋菌门显著增加(P<0.05);在属水平上,与对照组相比,亚麻籽组中norankf_Barnesiellaceae、GWE2-31-10、EMP-G18显著增加(P<0.05),短优杆菌群(Eubacterium_brachy_group)、norank_f_Chris-tensenellaceae显著减少(P<0.05)。
在属水平进行LEfSe多级差异物判别分析,并用LDA值衡量物种中差异效果影响大小。结果显示,对照组具有显著作用的菌有4个,分别为厌氧菌科(Anaerovoracaceae)、短优杆菌群(Eubacterium_brachy_group),未分类克里斯滕森氏菌属(norank_Christensenellaceae)、乳头杆菌属(Papillibact-er)。亚麻籽组具有显著作用的菌有12个,包括拟杆菌门下属科的理研菌科(Rikenellaceae)、巴恩斯氏菌科(Barnesiellaceae),norank_o Clostridia_vadinBB60_group、未分类巴恩斯氏菌科菌属(norank_Barnesiellaceae)及丹毒丝菌科(Erysipel-atoclostridiaceae)(图6)。
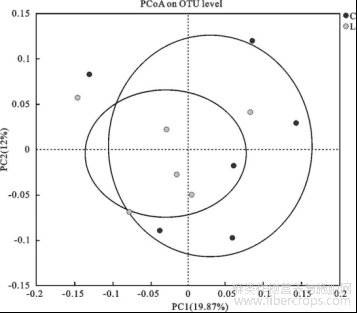
图2 亚麻籽对热应激西门塔尔牛粪便微生物β多样性的影响
A.门水平;B.属水平A.Atthe phylum level;B.Atthe genus level
图3 西门塔尔牛肠道菌群韦恩图
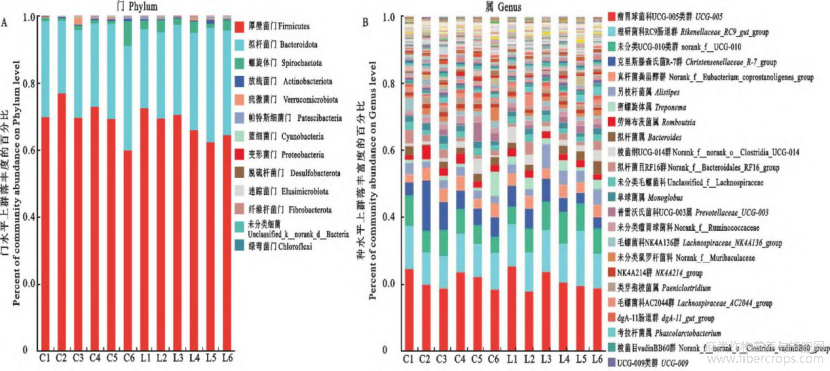
图4 西门塔尔牛肠道微生物在门(A)与属(B)水平的群落相对丰度分布
3 讨论
3.1 亚麻籽对热应激西门塔尔牛生理生化指标的影响
THI是评估动物热应激状态的核心环境参数,当THI>72时,牛即处于热应激状态。本试验期间牛舍THI平均值维持在78以上,且大部分时间处于中度热应激范围,为探究亚麻籽抗热应激作用提供了稳定的试验环境。公牛处于热应激状态时,睾丸及附睾温度升高,影响精子发生、精子活力、精子密度及总精子量[17]。公牛精子膜含有大量的PU-FAs,易受到LPO/ROS的侵害。亚麻籽含有亚油酸和α-亚麻酸等不饱和脂肪酸,是良好的抗氧化剂,其抗氧化特性能中和形成的LPO并改变膜流动性,降低质膜对ROS的敏感性,减少脂质过氧化产物的生成,从而缓解睾丸热应激,保护睾丸质膜免受LPO/ROS的侵害[18]。此外,油酸是亚麻籽油中存在的一种单不饱和脂肪酸,能够降低睾丸对脂质过氧化的易感性[2]。本试验中,与对照组相比,日粮中添加亚麻籽能够显著提升血清T-AOC、CAT的水平和降低MDA、COR含量,但对血清中SOD活性和HSP70水平影响不显著。Adeel等[19]对水牛补充亚麻籽油,提高了SOD、CAT、GSH和TAC抗氧化剂的生物利用度,降低了血清中MDA生成,与本试验结果一致,证明亚麻籽提高了动物体内抗氧化剂水平和不同抗氧化剂的生物利用度。热应激状态下机体下丘脑-垂体-肾上腺(HPA)轴激活,导致COR大量分泌,而过高的COR会抑制生殖激素合成、损伤睾丸生精功能。张道敬[20]研究表明,热应激肉鸡肾上腺分泌机能增加,甲状腺分泌机能降低,导致甲状腺和皮质激素分泌增加,血清中COR水平增加。本试验中亚麻籽组COR水平显著降低,表明亚麻籽可通过调节体内激素水平缓解公牛热应激。亚麻籽的抗氧化作用与其内在的抗氧化、ROS清除特性以及富含生物活性成分如omega-3脂肪酸和木脂素(lignan)有关,亚麻籽所含木脂素在消灭ROS和抑制ROS介导的DNA损伤方面具有很强的抗氧化和保护作用[21]。
3.2 亚麻籽对热应激西门塔尔牛精液品质的影响
精子质膜的完整性和结构稳定性是维持精子功能的基础,主要由磷脂和胆固醇构成,而磷脂中LC-PUFAs的含量直接影响膜的流动性、通透性及抗损伤能力。由于反刍动物自身无法合成ω-3PU-FAs,必须通过饲粮摄入,亚麻籽作为ω-3PUFAs的优质来源,其所含的α-亚麻酸可被机体吸收后,通过血液循环转运至睾丸组织,参与精子质膜的合成与修复[3]。在对种马[22]和火鸡[23]的研究中也发现饲粮补充omega-3脂肪酸对新鲜精液活力没有积极影响,热应激荷斯坦公牛膳食补充omega-3或其前体,可改善荷斯坦公牛新鲜精液的品质但对冻融精液品质的改善作用不明显[17]。本试验结果显示,亚麻籽组精子质膜完整率显著提高、畸形率显著降低。与此结论相似的是,公牛饲料补充亚麻籽油150mL/d可使精子功能膜完整性显著提高,精子在发情母牛宫颈黏液中向前推动的运动性、前进速度、质膜完整性和线粒体膜电位显著提高[2],兔子[3]和水牛[19]中也有类似结果。PUFAs对精子质膜完整性有显著影响,α-亚麻酸嵌入精子质膜后,优化了膜脂双分子层的组成比例,增强了质膜对氧化应激和渗透压变化的抵抗力,减少了热应激导致的质膜破裂。Kalo等[24]试验证明饲粮添加亚麻籽对精子密度和精子活力有积极作用,尼里拉菲水牛在基础饲粮水平上补饲120mL/d、250mL/d亚麻籽油,12周后精子密度、活力和质膜完整率显著提高。Yan等[25]研究表明,饲粮中添加n-3/n-6 PUFAs(比例为1.52∶1)和n-6/n-3(比例为1∶1)的大鼠睾丸和阴囊发育和精子形态结构更好,说明饲粮中添加适当比例的PUFAs促进了睾丸发育和精子发生。亚麻籽对精液品质的积极作用还与抗氧化剂有关,亚麻籽中的不饱和脂肪酸ω-3可保护精子尾部和中部的质膜免受脂质过氧化的影响,包括维生素E和酚类物质,这与雄性精子密度和运动能力的增强有关[5,26]。
3.3 亚麻籽对热应激西门塔尔牛粪便微生物菌群的影响
肠道微生物群作为机体“隐形器官”,通过参与营养物质代谢、调控免疫功能及合成生物活性物质,与宿主生殖健康密切相关,而热应激会破坏肠道菌群的平衡,导致有益菌丰度降低、有害菌增殖[27]。本试验中,α多样性分析显示两组肠道微生物的丰富度和多样性无显著差异,表明对照组和亚麻籽组粪便微生物多样性和丰富度相似,饲粮中添加亚麻籽对公牛肠道菌群没有产生明显改变。在组间显著性检验中,门水平上,与对照组相比,亚麻籽组中厚壁菌门显著减少,拟杆菌门和螺旋体门显著增加;厚壁菌门以分解纤维和淀粉为主,拟杆菌门则分解蛋白质和多糖,热应激状态下厚壁菌门丰度升高可能导致肠道发酵产物失衡,而亚麻籽的添加逆转了这一趋势,使拟杆菌门丰度增加[28]。Wang等[29]发现热应激会破坏公鸡肠道的稳定性,厚壁菌门相对丰度增加,拟杆菌门相对丰度减少。He等[30]指出热应激母猪阴道菌群中拟杆菌门的Bacteroidales RF16group_norank减少。而本试验发现添加亚麻籽后,厚壁菌门显著减少,拟杆菌门和螺旋体门显著增加,说明亚麻籽改善了公牛热应激状态下的菌群,使受热应激影响而失调的菌群回归正常比值。高温环境会影响肠道菌群数量、组成和代谢物质,高温环境下的黄羽公鸡盲肠中有益短链脂肪酸的拟杆菌(Bacteroidetes)相对丰度提高[31]。本试验发现对照组和亚麻籽组种丰度前三的菌属均含有拟杆菌门(Bacteroidota)。此外,分析对照组和亚麻籽组差异菌群,其中在门水平上,亚麻籽组中拟杆菌门显著增加。在属水平进行LEfSe多级差异物判别分析,对照组优势差异菌为厌氧菌科、短优杆菌属等,而亚麻籽组为理研菌科、巴恩斯氏菌科等。理研菌科作为拟杆菌门的下属科,其丰度增加与拟杆菌门的整体变化一致,已有研究表明理研菌科菌株可产生抗炎代谢产物,减轻肠道黏膜炎症,而肠道炎症的缓解可减少全身炎症反应对睾丸生精功能的抑制[31];巴恩斯氏菌科的丰度增加可能与营养物质代谢相关,其可能参与亚麻籽中不饱和脂肪酸的肠道代谢,促进ω-3PUFAs的吸收利用。相反,对照组中短优杆菌属丰度较高,该菌属部分菌株可能产生有害物质或加剧肠道菌群失衡,进而间接影响生殖功能[11]。Lin等[32]在公猪饲粮中添加膳食纤维增加了UCG-005和Rikenellaceae_RC9_gut_group等有益肠道微生物的丰度,降低了Romboutsia、Turicibacter等肠道有害微生物的相对丰度,促进短链脂肪酸的产生,从而提高了精子发生和精液质量。本试验证明热应激导致肠道菌群失衡,引发肠道代谢紊乱和炎症反应,同时热应激致使机体氧化应激,从而导致精液品质下降;而饲粮添加亚麻籽后,通过自身抗氧化成分直接缓解氧化应激,保护精子结构,还通过调节肠道菌群组成,优化肠道营养代谢,同时减少肠道炎症,从而达到缓解热应激的效果。
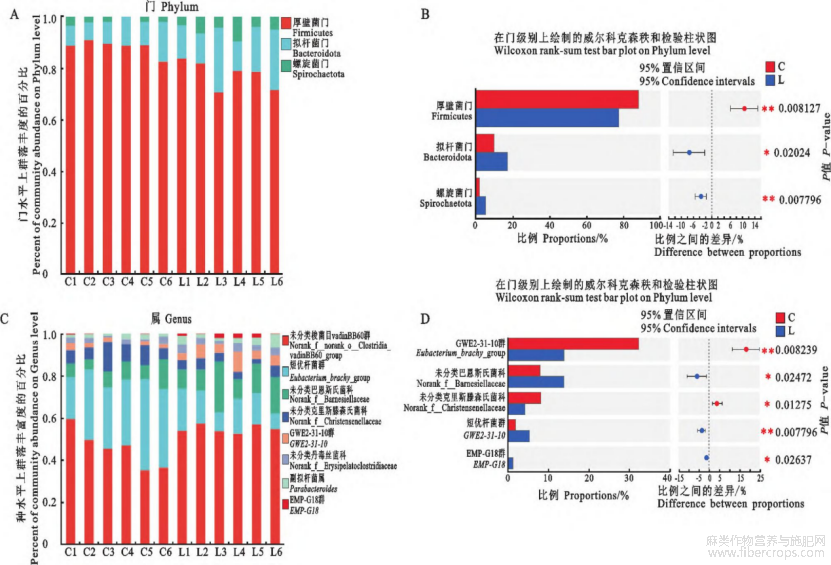
A.门水平的相对丰度分布;B.门水平组间差异菌群的Wilcoxon秩和检验分析;
C.属水平的相对丰度分布;D.属水平组间差异菌群的Wilcoxon秩和检验分析
图5 西门塔尔牛肠道微生物菌群的组成及组间差异分析
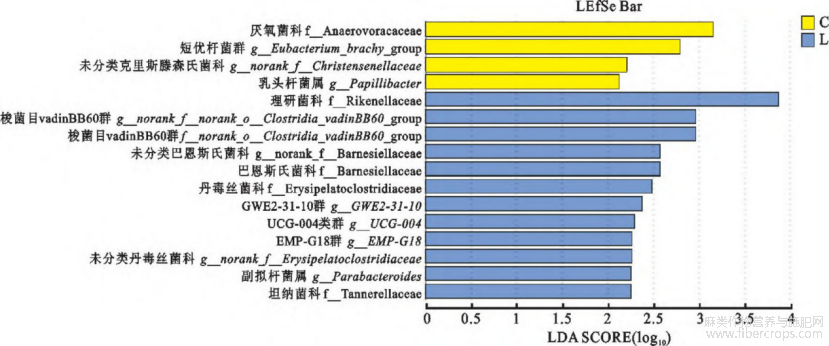
图6 对照组和亚麻籽组在属水平LDA判别结果表
4 结论
在热应激西门塔尔牛饲粮中添加亚麻籽可调节血清中的抗氧化酶水平,保护精子质膜免受ROS的侵害,提高精子顶体完整性,减少脂质过氧化,缓解热应激导致的精液品质下降。高温环境下肠道微生物群失衡会损害精子发生,西门塔尔牛饲粮添加亚麻籽后肠道中拟杆菌门丰度显著增加,厚壁菌门丰度减少,亚麻籽可能调节热应激公牛肠道微生物区系的相对丰度,提高机体对高温环境的耐受性,从而调控精液品质。
参考文献:
[1]王玉梅,杨拧嘉,霍晨曦,等.热应激对南阳牛种公牛精液品质的影响及对策[J].中国畜禽种业,2021,17(4):128-129.
[2]PERUMAL P,CHANG S,KHATEK,etal.Flaxseed oilmod-ulates semen production and its quality profiles,freezability,testicular biometricsand endocrinological profiles in mithun[J].Theriogenology,2019,136:47-59.
[3]MOURVAKIE,CARDINALIR,DALBOSCOA,etal.Effects offlaxseed dietary supplementationon sperm quality and on lipid composition of sperm subfractionsand prostatic granules in rabbit[J].Theriogenology,2010,73(5):629-637.
[4]MAHVIZ,MOGHADDASZADEH-AHRABIS,MOGADD-AMG.Effect of dietary Omega-3 and Lineseed oil on the sperm quality and fertility ofmale broiler breeders[J].Animal Science Research,2022,32(1):99-111.
[5]YUAN C,ZHANG K,WANG Z,etal.Dietary flaxseed oiland vitamin Eimprovesemen quality viapropionicacid metabolism [J].Frontiersin Endocrinology,2023,14:1139725.
[6]DE FRANÇA CARDOZO L F M,BOAVENTURA G T,BRANT L H C,etal.Prolonged consumption offlaxseed flour increases the 17β-estradiol hormone without causing adverse effects on the histomorphology ofWistar rats’penis[J].Food and chemicaltoxicology,2012,50(11):4092-4096.
[7]STANHISERJ,JUKICA,MCCONNAUGHEY D,etal.Ome-ga-3 fatty acid supplementation and fecundability[J].Human Reproduction,2022,37(5):1037-1046.
[8]FENG Y,DING Y,LIU J,et al.Effects of dietary omega-3/o-mega-6fatty acid ratioson reproduction in the young breeder rooster[J].BMC Veterinary Research,2015,11:73.
[9]SINGH M,MOLLIER R T,PONGENER N,et al.Linseed oil in boar’sdietduring high temperature humidity index (THI) period improves sperm quality characteristics, antioxidantsta-tusand fatty acid composition of sperm under hothumid sub-tropicalclimate[J].Theriogenology,2022,189:127-136.
[10]KIM SH,RAMOS SC,VALENCIA R A,et al.Heat stress:effects on rumen microbesand hostphysiology,and strategies to alleviate the negative impactson lactating dairy cows[J].Frontiersin microbiology,2022,13:804562.
[11]ZHU Y,LIX,SUOLANG,et al.House feeding pattern in-creased maleyak fertilitybyimprovinggutmicrobiotaand se-rum metabolites[J].Frontiersin Veterinary Science,2022,9:989908.
[12]WANG M,REN C,WANG P,et al.Microbiome-metabolome reveals thecontribution ofthegut-testisaxisto sperm motili-ty in sheep (Ovisaries)[J].Animals,2023,13(6):996.
[13]ASHONIBARE VJ,AKOREDE BA,ASHONIBARE PJ,et al.Gut microbiota-gonadal axis:the impact of gut microbiota on reproductive functions[J].Frontiersin Immunology,2024,15:1346035.
[14]NationalResearch CounciloftheNationalAcademies(NRC).Nutrient requirements of dairy cattle[S].Washington,D.C.:The NationalAcademies Press,2001.
[15]中华人民共和国农业农村部.牛冷冻精液:GB4143-2022[S].北京:中国标准出版社,2022.
[16]RAMU S,JEYENDRAN R S.The hypo-osmotic swelling testfor evaluation ofsperm membraneintegrity[J].Methods MolBiol,2013,927:21-25.
[17]GHOLAMIH,CHAMANIM,TOWHIDIA,et al.Improve-mentofsemen qualityin Holstein bulls during heatstress by dietary supplementation of omega-3 fatty acids[J].Interna-tionalJournalofFertility&Sterility,2011,4(4):160.
[18]ZANUSSI H P,SHARIATMADARIF,SHARAFIM,et al.Dietary supplementation withflaxseed oilassourceofomega-3 fatty acids improves seminal quality and reproductive per-formancein agedbroilerbreederroosters[J].Theriogenology,2019,130:41-48.
[19]ADEEL M,IJAZ A,ALEEM M,et al.Improvement of liquid and frozen-thawed semen quality of Nili-Ravi buffalo bulls (Bubalusbubalis) through supplementation offat[J].Ther-iogenology,2009,71(8):1220-1225.
[20]张道敬.不同抗热应激添加剂对肉鸡生产性能和血液理化指标的影响[J].南宁:广西大学,2002.
[21]NGCOBO JN,RAMUKHITHIFV,NEPHAWE K A,etal.Flaxseed oilasa source of omega n-3 fatty acids to improve semen quality from livestock animals:areview[J].Animals,2021,11(12):3395.
[22]BRINSKO S P,VARNER D D,LOVE C C,et al.Effect of feeding aDHA-enriched nutriceuticalon the quality offresh,cooled and frozen stallion semen[J].Theriogenology,2005,63(5):1519-1527.
[23]ZANIBONI L,RIZZI R,CEROLINI S.Combined effect of DHA and α-tocopherolenrichment on sperm quality and fer-tilityin the turkey[J].Theriogenology,2006,65(9):1813-1827.
[24]KALO D,RECHESD,NETTA N,et al.Carryover effects of feeding bulls with an omega-3-enriched-diet:From spermato-zoa to developed embryos[J].PLoSOne,2022,17(3):e0265650.
[25]YAN L,BAIXL,FANG Z F,etal.Effectofdifferentdietary omega-3/omega-6 fatty acid ratioson reproduction in male rats[J].Lipids in Health and Disease,2013,12:33.
[26]CARDOSO CARRARO JC,DANTASM IDS,ESPESCHIT A CR,et al.Flaxseed and huma-n health:reviewing benefits and adverse effects[J].Food Reviews International,2012,28(2):203-230.
[27]丁鸿祥,庞丽燕,王净,等.空怀绵羊粪便微生物和血液代谢组特征分析[J].动物营养学报.2022,34(5):3107-3121.
[28]SALES G FC,CARVALHO BF,SCHWAN RF,et al.Heat stressinfluencethemicrobiotaand organicacidsconcentration in beefcattlerumen[J].JournalofThermalBiology,2021,97:102897.
[29]WANG Y,SUN W,WU E,et al.Polysaccharides from Abrus cantoniensis hancemodulateintestinalmicrofloraand improve intestinalmucosalbarrier and liver oxidative damage induced by heat stress[J].Frontiers in Veterinary Science,2022,9:868433.
[30]HE J,ZHENG W,TAO C,et al.Heat stress during late ges-tation disrupts maternal microbial transmission with altered offspring’s gut microbial colonization and serum metabolites in a pig model[J].Environmental Pollution,2020,266:115111.
[31]张文彦,周倩,翟明仁.等.肠道菌群-胆汁酸”轴改善热应激畜禽肉品质的研究进展[J].动物营养学报,2024,36(1):55-63.
[32]LIN Y,WANG K,CHEL,et al.The improvement of semen quality by dietary fiber intake is positively related with gut microbiota and SCFA ina boar model[J].Frontiersin Micro-biology,2022,13:863315
文章摘自:徐逸飞,魏雅婷,李国才,等.亚麻籽对热应激西门塔尔牛生理生化、精液品质及后肠道菌群的影响[J].家畜生态学报,2026,47(04):46-55.
