摘要:为提高罗布麻生物脱胶效率,构建枯草芽孢杆菌与黄孢原毛平革菌的复合脱胶菌群,利用超临界CO2预处理破除罗布麻韧皮纤维的天然生物抗性屏障。结果表明:未经预处理的罗布麻在复合菌群脱胶后残胶率为19.6%,相较于单一菌种效果提升了26.9%;经超临界CO2预处理后脱胶,残胶率仅为10.8%。
关键词:罗布麻;生物脱胶;生物抗性屏障;超临界CO2;复合菌群
罗布麻属夹竹桃科多年生草本宿根植物[1],具有悠久的药用历史[2]。罗布麻被证实有良好的抗菌性[3-5],是一种极具潜力的天然功能性纤维原料。韧皮纤维制备中,微生物脱胶具有清洁性、可持续性的优势,但目前生物脱胶效率仍有待提高。就罗布麻生物脱胶而言,韧皮组织中由果胶、木质素和半纤维素等非纤维素物质构成的异质高聚物复合体和黄酮等天然抑菌物质,一起构成了罗布麻生物抗性屏障[6-8],这增加了脱胶过程的生物催化反应难度[9]。如何通过预处理破解生物抗性屏障,提高麻纤维生物的脱胶效率尤为重要。本研究针对罗布麻韧皮中非纤成分构建高效脱胶菌群,通过预处理破除韧皮的生物抗性屏障,从而提高生物脱胶效率。
1试验部分
1.1试验材料与仪器
罗布麻:新疆戈宝红麻有限公司。
菌种:枯草芽孢杆菌(天津工业大学清洁染整课题组Q)、黄孢原毛平革菌(广东省微生物菌种保藏中心GIM3.383)。
种子培养基:葡萄糖20.0g/L、蛋白胨10.0g/L、磷酸二氢钾3.0g/L、氯化钠2.0g/L、维生素B1微量。
发酵培养基:葡萄糖20.0g/L、磷酸二氢钾3.0g/L、硫酸镁1.5g/L、维生素B1微量。
试剂:葡萄糖(天津市光复科技发展有限公司),蛋白胨(北京普博欣生物试剂公司),硫酸镁(天津大学科威公司),磷酸二氢钾(天津市化学试剂六厂),无水乙醇(天津市江天化工有限公司),维生素B1、3,5-二硝基水杨酸(天津光复精细化工有限公司),酒石酸钾钠、苯酚、氢氧化钠(天津科密欧化学试剂有限公司)。
仪器:电子分析天平(天津天马衡基仪器有限公司),电热鼓风干燥箱(天津市中环实验电炉有限公司),单人单面净化工作台(苏州净化设备有限公司),生化培养箱(上海新苗医疗器械制造有限公司),立式压力蒸汽灭菌器(上海申安医疗器械厂),紫外可见分光光度计(北京普析通用仪器有限责任公司),台式扫描电镜,傅立叶变换量红外光谱仪(美国赛默飞世尔科技),全自动物理化学吸附仪(美国康塔公司),X射线衍射仪(德国BRUKER公司)。
1.2试验方法
1.2.1超临界CO2预处理
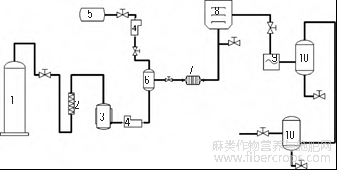
将罗布麻韧皮纤维在50℃中烘至干燥,干燥后的罗布麻韧皮纤维剪成5cm长短的样品,用医用纱布包裹放入超临界发生装置(如图1所示)的反应釜中,无水乙醇为夹带剂。处理温度45℃,压力20MPa,总流量30.0g/L,夹带剂含2%,处理60min。
1. CO2气瓶;2.净化器;3.制冷机;4.高压泵;5.夹带剂储罐;6.混合器;7.热交换器;8.反应釜;9.变压装置;10.分离釜
图1 超临界CO2预处理装置示意图
1.2.2生物量测试
枯草芽孢杆菌为细菌,生物量测试选用600nm波长进行光电比浊测定,用未接种的液体培养基作空白对照。枯草芽孢杆菌按5%的接种量接种到培养基中,每隔4h取样测定,测定前将培养液充分振荡,使微生物分布均匀。以光密度值为纵坐标,培养时间为横坐标绘制枯草芽孢杆菌的生长曲线。
黄孢原毛平革菌是真菌,生物量测试采用菌丝干燥质量法,以菌体干燥质量为纵坐标,培养时间为横坐标,绘制黄孢原毛平革菌生长曲线。
1.2.3酶活测试
果胶酶酶活测试采用DNS法,漆酶活性的测定采用ABTS法[10]。
1.2.4脱胶效果表征
用质量损失率表征脱胶效果,计算方法见式(1)。
![]()
式中:w为质量损失率,%;w0为脱胶前样品质量,g;w1为脱胶后样品质量,g。按照GB5889—86《苎麻化学成分定量分析方法》对罗布麻纤维进行残胶率测定,计算方法见式(2)。
![]()
式中:ω为残胶率,%;m0为试验前罗布麻韧皮纤维质量,g;m1为试验后韧皮纤维质量;g。
2结果与讨论
2.1罗布麻韧皮的超临界CO2处理
高比例非纤维素物质、致密多层网状结构和黄酮等抑菌物质成分构成了罗布麻韧皮纤维生物抗性屏障[4,9]。为提高生物可及性,用超临界二氧化碳对罗布麻纤维进行预处理研究。
2.1.1形态结构
观察预处理前后罗布麻韧皮纤维形态结构的变化,结果如图2所示。

(a)罗布麻原麻

(b)超临界CO2处理后的纤维
图2 预处理前后罗布麻韧皮纤维电镜图(×500)
由图2可知,罗布麻韧皮纤维是束纤维,其中单纤维被胶质层紧密包裹。束纤维外的胶质层在脱胶过程阻碍微生物及酶与底物接触,是生物脱胶的第一层屏障。预处理后,纤维结构松散开来,单纤维间的距离增大,且包裹在纤维上的胶减少。这说明预处理不仅可以改变罗布麻韧皮纤维的形态结构,同时还能去除部分胶质,破坏纤维内致密的网状结构,提高生物催化剂的可及性。
罗布麻韧皮组织内部的孔隙结构对酶与底物的接触有重要影响。为考察预处理前后罗布麻韧皮组织内部孔隙结构的变化,利用全自动物理化学吸附仪,对样品的比表面积、孔容和孔径进行测试,结果如图3所示。
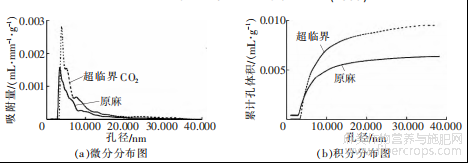
图3 预处理前后罗布麻韧皮纤维孔径分布图
孔径分布代表着韧皮纤维内部微细孔的容积随孔径尺寸的变化情况,是表征物质孔隙结构的重要参数之一。由图3中的数据可以看出,罗布麻韧皮纤维处理前后孔径在1.000~40.000nm之间,在3.000~15.000nm分布最广。预处理后,3.000~15.000nm孔径明显增加,而且累计孔体积增加显著。这说明超临界CO2处理能有效地改善罗布麻纤维内部孔隙结构,提高韧皮纤维内部的孔径和孔容。
计算罗布麻韧皮纤维处理前后比表面积和体积,结果见表1。
表1 预处理前后罗布麻韧皮纤维孔隙结构
|
项目 |
孔径/ nm |
比表面积/ (m2·g-1) |
体积/ (mL·g-1) |
|
原麻 |
3.433 |
6.545 |
0.006 |
|
超临界CO2 |
3.806 |
6.036 |
0.010 |
由表1可知,超临界预处理后罗布麻韧皮纤维比表面积小幅度减小,孔径和体积显著增大。这为生物大分子进入纤维内提供了更多空间,有利于生物脱胶的进行。
2.1.2超分子结构
为进一步研究预处理对罗布麻韧皮纤维结构的影响,测试了预处理前后样品的超分子结构,结果如图4所示。
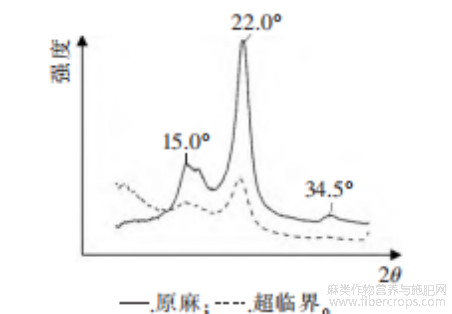
图4 预处理前后罗布麻韧皮纤维XRD图谱
由图4可知,超临界CO2预处理后的罗布麻韧皮纤维XRD衍射图谱在2θ=5.0°~13.0°出现明显的变化,101晶面(2θ=15.0°)、200晶面(2θ=22.0°)和040晶面(2θ=34.5°)说明了罗布麻韧皮纤维在预处理前后都属于典型纤维素Ⅰ晶型[11]。由预处理后101晶面、200晶面和040晶面的衍射峰,可以看出处理后麻的晶态结构有明显变化
计算处理前后罗布麻韧皮纤维的晶面间距和晶粒尺寸,见表2。
|
项目 |
结晶 度/% |
晶面间 距/nm |
晶粒尺 寸/nm |
|
原麻 |
74 |
4.73 |
2.22 |
|
超临界CO2 |
52 |
6.65 |
2.12 |
表2 预处理前后罗布麻韧皮纤维结晶参数
该结果表明,超临界CO2处理后结晶度下降幅度大,达到了22%,晶面间距明显增加。在超临界CO2处理中,CO2渗透进罗布麻韧皮纤维中,使其发生溶胀,分子间的距离增大,分子间的相互作用减小[12],可降低纤维素Ⅰ晶型结晶时的能垒,破坏结晶区的纤维素,改变各项结晶参数[13]。结晶度高的纤维溶胀比较困难[14],结晶度下降,无定形的纤维素分子间和分子内的氢键作用力减弱,分子链上的羟基大部分暴露出来,增强了纤维素和木质素的可及度和反应活性,也增强了脱胶过程中水的溶胀作用;韧皮纤维的晶面结构被破坏,有利于微生物和酶渗透进入纤维内部与木质素反应,提高脱胶效率。
2.1.3分子结构
研究预处理对罗布麻韧皮纤维结构组成的影响,用傅立叶变换红外光谱仪对预处理前后的样品进行测试,结果如图5所示。

图5 预处理前后罗布麻韧皮纤维红外光谱图
图5显示了罗布麻韧皮纤维超临界CO2处理前后的分子结构变化。结果表明,处理前后的样品在3420cm-1、1432cm-1、1164cm-1和1058cm-1处均有吸收峰,3420cm-1的吸收峰属于羟基O—H伸缩振动,主要代表纤维素,而1432cm-1、1164cm-1和1058cm-1则是纤维素结构在指纹区的吸收峰。
3337cm-1和3283cm-1处是O—H伸缩振动,超临界CO2处理后,此处吸收峰峰强减弱,说明处理过程伴随着纤维素的损失。
2920cm-1、2848cm-1处的吸收峰是蜡质的饱和烷烃C—H的拉伸振动[15]。处理后2850cm-1处的吸收峰消失,表明过程中蜡质被去除。蜡质层有很强的疏水性,处理过程中,超临界CO2流体由于分子较小可以渗透进入罗布麻韧皮纤维内部,从而破坏其生物抗性屏障,相当一部分的胶质在此过程中可以被破碎带出,增强了后续试验过程中水的溶胀作用。
1737cm-1处的振动峰,是果胶中的甲酯和羧酸或半纤维素中的乙酰基的C=O伸缩引起的,经过超临界CO2处理的样品光谱在此处的吸收峰接近消失,说明超临界CO2处理对果胶和半纤维素的去除效果优异。
1599cm-1为C=O和C=C伸缩振动的叠加,为韧皮中黄酮提取物的特征峰。超临界CO2处理后,此处吸收峰有了显著减弱,说明超临界CO2能萃取出韧皮中的黄酮物质,削弱其抑菌性,有利于微生物渗透生长。
1428cm-1和1463cm-1是芳香族骨架振动和C—H的拉伸振动,与罗布麻韧皮纤维中的木质素有关。超临界CO2处理后此处的吸收峰都有大幅度降低,表明超临界CO2能够有效地去除韧皮中的木质素。
罗布麻韧皮纤维在超临界CO2处理过程中,胶质成分的去除,减轻了后续生物脱胶的负担。同时,由于大量非纤成分的去除,使得罗布麻韧皮组织结构变得疏松,增大了生物催化剂的可及度。
2.2罗布麻高效脱胶菌群的构建
脱胶菌种是生物脱胶最核心的部分,罗布麻相较于苎麻等麻类纤维具有更高的木质素含量,这给生物脱胶带来了很大的阻碍。以往对罗布麻生物脱胶的研究大多使用细菌,如枯草芽孢杆菌、地衣芽孢杆菌等芽孢杆菌[6-7,16-17]。随着纺织生物技术研究的不断深入,大量真菌如白腐菌、褐腐菌、软腐菌也被发现能降解木质素,其中白腐菌是已知唯一能够将木质素彻底降解为二氧化碳和水的生物,已被广泛应用于造纸行业和生物炼制[18-22]。
微生物菌落的生物多样性对生态系统功能的影响,已被试验证实可以通过选择特定的微生物组合来设计菌落,以获得最大的生态系统功能速率[23]。本试验尝试将黄孢原毛平革菌与枯草芽孢杆菌进行混合培养,考察它们的产酶能力和脱胶能力,建立高效复合脱胶菌群。
2.2.1菌群的生长特性
枯草芽孢杆菌按5%接种量接种,黄孢原毛平革菌按10%接种量接种,以1:2的接种量构建复合菌群,按1.2.2方法分别测试不同生长时期单一枯草芽孢杆菌和黄孢原毛平革菌的生物量以及复合菌群中它们的生长情况,绘制生长曲线如图6所示。
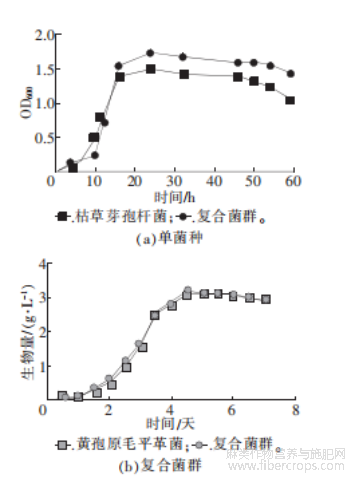
图6 单菌种与复合菌群生长情况图
由图6a可以看出,枯草芽孢杆菌在开始培养的前10h内,生物量呈缓慢上升状态;10h至17h生物量急剧增长,菌体进入对数生长期;45h后,由于营养物质的匮乏导致菌体开始衰亡,生物量呈下降趋势。与黄孢原毛平革菌组成的复合菌群生长曲线趋势和单一枯草芽孢杆菌类似,其进入对数生长期的时间更早,培养6h后生物量开始急剧增长,有更久的稳定生长期,且相同时间生物量远超单一枯草芽孢杆菌,因此在该菌落中,枯草芽孢杆菌拥有更适宜的生长环境。
图6b显示,黄孢原毛平革菌从第2天开始进入对数生长期,第5天时稳定生长,第7天开始进入衰亡。不同时间段复合菌群的生物量相较于单一黄孢原毛平革菌略有提高,两者整体生长情况近似,表明复合菌群的环境对黄孢原毛平革菌的生长没有太大影响。
2.2.2菌群的酶活
大量研究表明,本试验所用枯草芽孢杆菌主要产果胶酶、木聚糖酶和葡甘露聚糖酶,其中果胶酶酶活最高[6]。黄孢原毛平革菌是白腐真菌的一种,主要分泌漆酶(LaC)、木质素过氧化物酶(LiP)和锰过氧化物酶(MnP),其中LiP和MnP的合成条件较为苛刻,氮源浓度、Mn2+浓度及含氧量都会对它们产生显著的影响[24]。因此,为比较复合菌群与单一菌种的代谢能力,按1.2.3分别测试枯草芽孢杆菌果胶酶酶活、黄孢原毛平革菌漆酶酶活以及复合菌群的果胶酶和漆酶酶活,结果如图7所示。
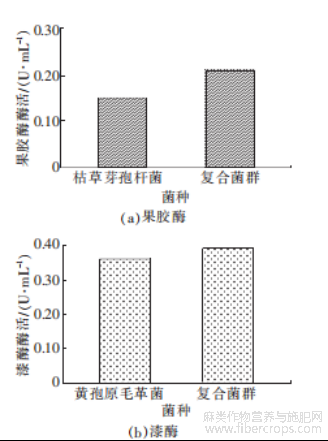
图7 菌群的酶活
由图7可知,枯草芽孢杆菌在生长过程中分泌的果胶酶酶活为0.15U/mL,黄孢原毛平革菌产的漆酶酶活为0.36U/mL,而在复合菌群中,果胶酶酶活达到0.21U/mL,提升了40.00%,漆酶酶活同样也提升了8.33%。在这样一个复合脱胶菌落中,产漆酶的能力强于产果胶酶,表明该菌群对木质素具有更好的降解效果,复合菌群中两种酶的酶活相较于单一菌更强,证明该菌落对两种菌的生长代谢都有促进作用,可以用于生物脱胶。
2.2.3菌群的脱胶能力
为表征各脱胶菌种(群)脱胶能力,参考1.2.4方法分别测试枯草芽孢杆菌、黄孢原毛平革菌和复合菌群的脱胶能力,如图8所示。
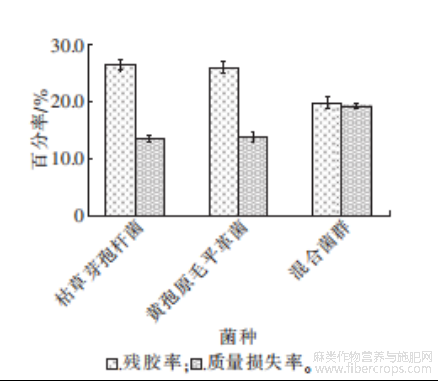
图8 不同菌种脱胶效果对比图
从图8可以看出,使用单一菌种对罗布麻韧皮纤维进行脱胶时,残胶率都在26.0%,质量损失率仅有14.0%~15.0%。枯草芽孢杆菌主要产果胶酶和木聚糖酶,脱胶过程中主要针对果胶、半纤维素以及少量的木质素,黄孢原毛平革菌产漆酶(LaC)、木质素过氧化物酶(LiP)和锰过氧化物酶(MnP),全部用于降解木质素,因此使用单一菌种脱胶时容易造成脱胶不彻底。使用由枯草芽孢杆菌和黄孢原毛平革菌构成的复合脱胶菌群脱胶时,质量损失率上升至19.1%,残胶率下降为19.6%,效果较单一菌种脱胶时提升26.9%,复合菌群中微生物的协同作用可以促进生物脱胶过程的高效运行。然而罗布麻韧皮纤维生物抗性屏障的存在,使得脱胶效果仍不理想。
2.3抗性屏障破除条件下高效菌群的生物脱胶
为探究罗布麻韧皮纤维抗性屏障的破除对高效脱胶菌群的影响,从菌群的生长情况和脱胶能力两个方面进行比较。
2.3.1抗性屏障破除条件下高效菌群生长情况
罗布麻韧皮纤维中含有黄酮、鞣质等抑菌物质,在脱胶过程中不利于微生物生长繁殖,为比较超临界CO2处理对这类抑菌物质的影响,将预处理前后的罗布麻韧皮纤维粉末作为主要营养物质培养高效复合菌群,按1.2.2测试不同阶段的生物量,绘制生长曲线如图9所示。
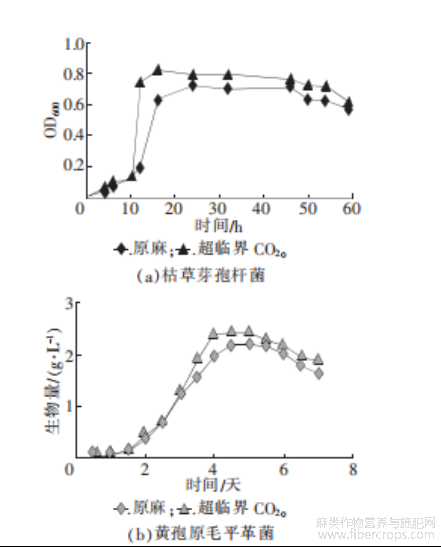
图9 超临界CO2处理前后菌群生长情况
由图9可知,经过预处理后菌群的生物量相较于不做处理的原麻显著提升。在用超临界CO2进行处理时,不仅罗布麻韧皮纤维内部的形态结构发生了变化,以此提高了生物催化剂的可及度,使微生物更容易进入到纤维内部,获得更多的营养物质进行生长繁殖,它的分子结构同样发生了改变,黄酮等抑菌物质在此过程中被萃取出来,此时的罗布麻韧皮更适宜微生物的繁殖,因此菌株的生长更为迅速,在后续脱胶过程中更有利于脱胶菌的生长繁殖。
2.3.2抗性屏障破除条件下高效菌群脱胶效果
为探究抗性屏障对高效菌群脱胶能力的影响,对超临界CO2处理前后的罗布麻韧皮纤维用高效菌群进行脱胶,参考1.2.4的方法进行表征,结果如图10所示。
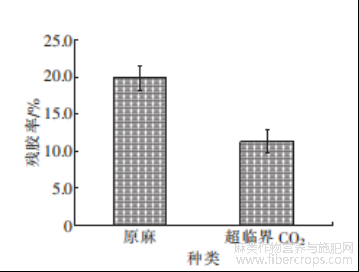
图10 超临界CO2处理前后脱胶效果对比图
由图10可知,未经预处理的罗布麻原麻直接进行脱胶,残胶率较高,经过超临界CO2处理后脱胶,残胶率为10.8%,效果显著提升。由于罗布麻韧皮纤维天然生物25.0抗性屏障的存在,使得脱胶菌很难渗透进纤维内部,导致脱胶不彻底,且纤维内部的抑菌物质也会抑制微生物生长,不利于生物脱胶;超临界CO2预处理不仅可以利用CO2气体使纤维内部膨胀松散开,改变韧皮内部的孔隙结构,破除生物抗性屏障,提高生物催化剂的可及度,在此过程中还伴随着果胶、木质素的少量去除[25],减轻后续生物脱胶的压力,还能萃取出相当的黄酮等抑菌物质,有利于脱胶菌的生长代谢,提高脱胶效率。
3结论
3.1超临界CO2处理能有效地破除罗布麻韧皮纤维的抗性屏障,萃取出纤维内的抑菌物质,使得纤维孔径增大,结晶度降低,提高生物催化剂的可及度。
3.2复合菌群对枯草芽孢杆菌和黄孢原毛平革菌的生长和产酶能力都有促进作用,复合菌群的脱胶能力也优于任意单一菌种。
3.3生物抗性屏障被破除后的罗布麻更适合高效复合脱胶菌群的生长繁殖,对纤维进行处理后脱胶,脱胶效果提升8.0%~9.0%。
参考文献
[1]张卫明,肖正春,张广伦,等.新疆罗布麻生态类型及其纤维品质研究[J].中国野生植物资源,2006(4):33-37.
[2]GRUNDMANN O, NAKAJIMA J I,KAMATA K, et al. Kaempferol from the leaves of apocynum venetum possesses anxiolytic activities in the elevated plus maze test in mice [ J ] . Phytomedicine , 2009,16(4):295-302.
[3]LI M, HAN G, CHEN H, et al. Chemical compounds and antimicrobial activity of volatile oils from bast and fibers of apocynum venetum [J].Fibers and Polymers,2012,13(3):322-328.
[4]XU X, GONG J, ZHANG T, et al. Insights into antibacterial mechanism of apocynum venetum L. fiber: evolution of bioactive natural substances in bast dur ing chemical degumming process [J].In- dustrial Crops and Products,2020(151):112419.
[5]李明华.罗布麻纤维抑菌成分与抑菌性能的研究[D].上海:东华大学,2011.
[6]张涛.基于抗性屏障结构破除的罗布麻纤维生物脱胶研究[D].天津:天津工业大学,2019.
[7]张秋亚.罗布麻纤维仿生集成脱胶研究[D].天津:天津工业大学,2017.
[8]WANG L, HAN G, ZHANG Y. Com-parative study of composition, structure and properties of apocynum venetum fibers under different pretreatments [J]. Carbohydrate Polymers,2007,69(2):391-397.
[9]巩继贤,张秋亚,张涛,等.韧皮结构对罗布麻生物脱胶的影响[J].纺织学报,2017,38(12):83-87.
[10]PAMIDIPATI S, AHMED A. A first report on competitive inhibition of lac- case enzyme by lignin degradation in- termediates[J].Folia Microbiol (Praha),2020,65(2):431-437.
[11]王磊磊.罗布麻纤维结构及成分初探[D].青岛:青岛大学,2007.
[12]刘颋,曹贵平,刘涛,等.高压CO2对聚丙烯溶胀行为的影响[J].高分子材料科学与工程,2005(6):130-133.
[13]高世会.罗布麻韧皮纤维超临界CO2协同生物化学脱胶研究[D].上海:东华大学,2016.
[14]朱国华,俞建勇.亚麻脱胶后纤维的纵向结构和强力的研究[J].上海纺织科技,2004(3):11-12.
[15]LOJEWSK J, MISKOWIEC P,LOJEWSKI T, et al. Cellulose oxidative and hydrolytic degradation: in situ FTIR approach[J].Polymer Degradation & Stability,2005,88(3):512-520.
[16]郑权莉,巩继贤,李辉芹,等.微生物处理条件下罗布麻脱胶动态过程分析[J].针织工业,2015(1):41-44.
[17]杨琦,段盛文,彭源德.苎麻微生物脱胶技术的研究进展[J].中国麻业科学,2018,40(1):36-42.
[18]朱丽平.白腐菌高效降解竹材木质素的工艺条件研究[D].重庆:重庆大学,2009.
[19]龚国利,陈志宣,陈松,等.高产漆酶白腐真菌的分离与鉴别[J].陕西科技大学学报:自然科学版,2012,30(6):79-83.
[20]SHINDE S D, MENG X, KUMARR, et al. Recent advances in under- standing the pseudo -lignin formation in a lignocellulosic biorefinery [ J ]. Green Chemistry,2018,20(10):2192-2205.
[21]张超,张晓晖,王家玺,等.木质素生物炼制研究进展[J].氨基酸和生物资源,2016,38(4):60-70.
[22]林海龙.木质纤维素生物炼制的研究进展[J].生物加工过程,2017,15(6):44-54.
[23]BELL T, NEWMAN J A, SILVER-MAN B W, et al. The contribution of species richness and composition to bac terial services[J].Nature,2005,436(7054):1157-1160.
[24]李华钟,章燕芳,华兆哲,等.黄孢原毛平革菌选择性合成木质素过氧化物酶和锰过氧化物酶[J].过程工程学报,2002(2):137-141.
[25]SEGHINI M C, TOUCHARD F,CHOCINSKI-ARNAUIT L, et al. Envi- ronmentally friendly surface modification treatment of flax fibers by supercritical carbon dioxide[J].Molecules(Basel, Swit- zerland),2020,25(3):438.
文章摘自:罗子都,任远,李政,巩继贤基于抗性屏障破除的罗布麻高效菌群脱胶研究针织工业[J]2022年第6期
