摘 要:【目的】为探究两种生态型大麻杂交F2代群体在不同生态环境种植下主要农艺性状间的遗传变异特点。【方法】以‘云麻1号’和‘Dame Blanche’杂交获得的F2代株系群体作为研究对象,在3种生态环境种植下对亲本和F2代分离群体的株高、茎粗和分枝数等10个农艺性状进行测定,并对这些农艺性状进行遗传变异分析、相关性分析及广义遗传力估算。【结果】8个农艺性状在 3种生态环境种植下表现出显著差异(P<0.05)。F2代群体中多数农艺性状平均值介于双亲之间,但群体中各农艺性状变异丰富,在3个生态环境中,10个农艺性状变异系数均超过10%,平均变异系数(3个生态环境下变异系数之和/3)由大到小依次为:第一分枝高>鲜重>分枝长>分枝数>株高>茎粗>叶宽>小叶宽>叶长宽比>叶长。各农艺性状间关系密切,除小叶宽、叶宽、叶长宽比(叶形)3个性状外,其余7个农艺性状在3个生态环境下均两两呈极显著正相关性。株高、分枝数、分枝长等农艺性状分布呈连续正态分布,具有典型的数量性状遗传特征。10个农艺性状广义遗传力变化幅度为29.508%~82.575%,由大到小依次为:分枝长>株高>第一分枝高>叶长>分枝数>茎粗>叶宽>小叶宽>鲜重>叶长宽比,其中分枝长和株高广义遗传力超过80%。【结论】在F2代群体中10个农艺性状变异丰富,表现出广泛的性状分离,各农艺性状间关系密切,具有典型的数量性状遗传特征。本研究初步探讨了两种生态型大麻杂交F2代主要农艺性状遗传分离特征,结论可为大麻不同用途品种选育和相关性状的QTL定位提供理论基础。
关键词:大麻;农艺性状;杂交F2代;遗传变异;广义遗传力
【研究意义】大麻(Cannabis sativa L.)为大麻科大麻属一年生草本植物,多雌雄异株[1]。大麻植物学分类还存在一定争议,有学者认为大麻至少存在indica和sativa两个亚种或变种(或生态型)[2–4],二者形态差异较大,前者普遍株型矮小,节间较短,分枝紧凑,后者则株型高大,节间较长,分枝稀疏。作为一种传统的经济作物,大麻各种用途都与农艺性状有密切关系,但长期以来由于中国工业大麻品种(指花、叶四氢大麻酚含量<0.3%,无毒品利用价值的大麻品种)选育亲本多来自国内种质资源,亲本来源范围小,且大多数属于sativa生态型,使得工业大麻品种同质化较严重,遗传基础趋同[5]。此外,由于大麻为短日照植物,易受光照、温度、土壤等环境因素影响[6–7],导致品种适应范围有限。因此,育种工作者想通过扩大亲本来源,选育出能满足不同用途和栽培环境的品种。【前人研究进展】株高、茎粗等表型性状与大麻的生物产量和收获指数有密切联系[8],培育具有优良农艺性状的新品种是提高大麻产量及品质的有效方法之一。长期的生产实践表明,纤维用大麻植株以节数少、分枝少、株高和茎粗适中及叶少为优[9];而以籽用为主时,良好的株型结构能提高光合利用率[10],进而提高麻籽产量,在一定范围内,籽粒产量与株高、茎粗、分枝数等农艺性状存在正相关性[11];以药用为主时,因药用成分(大麻素,Cannabinoid)主要存在于雌株花叶中[12–13],花叶产量又与分枝数、株高等农艺性状有密切关系。汤志成等[14]通过对16份大麻种质资源(其中12份为野生种质资源,4份为栽培品种)表型性状进行遗传多样性分析,结果表明中国野生大麻种质资源表型变异大,遗传多样性丰富。吕咏梅等[15]对大麻不同时期7个农艺性状进行了广义遗传力分析,结果表明大麻工艺成熟期株高、茎粗、鲜重等农艺性状广义遗传力较快速生长期要大。Petit等[16]利用123份大麻种质材料开展了与纤维品质有关的28个农艺性状的遗传变异研究,结果表明株高、茎粗和花期等农艺性状在不同生态环境下差异显著。【本研究切入点】农艺性状对大麻栽培生产和目标产物的获得有极大影响。目前对大麻不同亚种杂交后代农艺性状进行遗传变异及相关性研究鲜有报道,尚没有对大麻分离群体在不同生态环境下开展农艺性状遗传的研究。尽管获得优良农艺性状的方法很多,但杂交育种方法仍是最常见且最有效的方法之一。陈璇等[5]认为在工业大麻品种选育过程中,应扩大品种的遗传背景,尽可能选择地理分布较远,目标性状差异较大的亲本,避免品种同质化现象。【拟解决的关键问题】因此,本研究选用两个农艺性状差异较大且分属于不同生态型的大麻材料作为亲本,通过化学诱导构建F2代全雌性株系群体,将F2代株系群体种植于3个不同生态环境下,对每个生态环境下植株的株高、茎粗、叶长宽比(叶形)等10个农艺性状进行测定,并对各农艺性状进行变异分析、相关性分析及遗传力分析,探究大麻F2代株系群体在不同生态环境种植下各农艺性状间的关系及遗传特点,以期为大麻不同用途品种选育及相关性状的QTL定位提供理论参考。
1材料与方法
1.1试验地点
2018年7—10月在云南省玉溪市峨山县化念镇、昆明市嵩明县小街镇和昭通市巧家县马树镇3个生态环境下开展F2代群体多点种植评价试验。峨山县化念镇种植点经纬度为东经102°19′,北纬24°07′,海拔1120m,年平均气温20℃,年平均降雨量828mm,土质为沙壤土;嵩明县小街镇种植点经纬度为东经103º12′,北纬25º30′,海拔1910m,年平均气温14.1℃,年平均降雨量996mm,土质为红壤土;巧家县马树镇种植点经纬度为东经103º25′,北纬26º75′,海拔2420m,年平均温度8℃,年平均降雨量726.4mm,土质为红壤土。
1.2供试材料
本研究所用亲本材料为中国第一个工业大麻品种‘云麻1号’(简称YM1)和国外引进品种‘Dame Blanche’(简称Dame),将2份亲本材料种植于昆明市盘龙区进行表型特性观测统计(表1)。利用化学诱导性别转换技术(专利号:ZL201410506082.7)开展了两份材料的自交纯合,获得可用于F2代株系群体构建的两份相对纯合自交2代亲本材料。
表1 两份大麻亲本材料的表型特性

1.3 F2代全雌性株系群体构建
从YM1与Dame自交2代材料中各选取1株雌株作为杂交亲本,其中YM1雌株经化学诱导使其开雄花,授粉Dame雌株,收籽得到F1群体。选取F1群体中一个单株进行扦插繁殖得到一个克隆群体,化学诱导部分植株开雄花,授粉收籽即得到全雌F2群体。F2播种后,随机选取300个单株进行编号,并将每一个单株扦插培育成株系,所有株系同时移栽至峨山县、嵩明县和巧家县3个生态环境下,每个生态环境下每个株系确保3株克隆株(重复)。种植株行距为1.0m×1.6m,统一施肥、除草等田间管理。
1.4农艺性状测定及广义遗传力估算
参照大麻种质资源描述规范和数据标准[17]并结合本研究实际情况,于盛花末期对F2代株系群体中植株的株高、茎粗、第一分枝高、分枝数、分枝长、叶长(复叶)、叶宽(复叶)、叶长宽比(复叶)、小叶宽、鲜重共10个农艺性状进行数据统计。其中,分枝长为植株中部最长两个分枝的平均值,小叶宽为复叶最中间一片小叶的宽度。并按照以下公式对各性状的广义遗传力进行估算[18]。

1.5数据处理
使用Excel 2016软件对试验数据进行描述统计和图表绘制,使用SPSS 26.0软件进行方差分析、相关性分析和均方计算。
2结果与分析
2.1双亲和F2代农艺性状比较
F2代株系群体在3个生态环境种植下,除叶长在嵩明和巧家地区没有表现出显著差异(P<0.05)外,其它农艺性状在3个生态环境下均表现出显著差异。其中,株高、第一分枝高、分枝数、分枝长和叶长宽比5个农艺性状平均值在嵩明地区种植的F2代株系群体中最大;茎粗、叶长、叶宽和小叶宽4个农艺性状平均值则在巧家地区种植的F2代株系群体中最大;而在峨山地区种植的F2代株系群体中,所有农艺性状平均值均为最小(表2)。
在3个生态环境种植下的F2代株系群体中,各性状最大值均大于高值亲本,最小值均小于低值亲本(表3),表现出广泛的性状分离,丰富的性状分离使群体中大部分性状平均值最终居于双亲之间。而在峨山地区种植的F2代群体中,叶长宽比平均值则小于低值亲本;在巧家地区种植的F2代群体中,分枝数平均值大于高值亲本;3个生态环境下,小叶宽趋向或低于低值亲本。
表2 大麻亲本及其F2代农艺性状数据比较

注:表中数值为平均值±标准差,F2中同一性状不同小写字母表示在不同生态环境下差异显著(P<0.05),“-”表示数据缺失。
2.2 F2代农艺性状变异分析
通过对3个生态环境下F2代株系群体中10个农艺性状进行变异分析,结果表明:10个农艺性状变异程度差异较大,变异系数均大于10%,变异幅度为13.12%~74.64%(表3),10个农艺性状平均变异系数(3个生态环境下变异系数之和/3)由大到小依次为:第一分枝高>鲜重>分枝长>分枝数>株高>茎粗>叶宽>小叶宽>叶长宽比>叶长。10个农艺性状中,平均变异系数最小的为叶长,但其变异系数也达到了17.79%,表明这10个农艺性状通过杂交选择改良潜力较大。
某些性状的变异程度在不同生态环境下表现出较大差异。峨山地区除鲜重数据缺失外,其余农艺性状变异系数均大于另外2个地区。第一分枝高、分枝数、分枝长3个与分枝有关的农艺性状在不同地区变异系数差异较大,分枝数在峨山和巧家2个生态环境下变异系数相差近50%,第一分枝高也相差近39%,而其它7个农艺性状在不同生态环境下变异系数差异则相对较小。以上结果表明,对大麻不同农艺性状进行选择时要考虑其在特定环境中的变异程度,并根据不同的生态环境选育适应的品种。
表3 F2代株系群体10个农艺性状遗传变异比较

2.3 F2代农艺性状相关性分析
对在3个生态环境种植下的F2代群体中10个农艺性状进行相关性分析。由表4-表6可知,3个生态环境下,株高、茎粗、第一分枝高、分枝数、分枝长、叶长6个农艺性状间均呈显著或极显著正相关关系(表4-表6)。其中,株高与第一分枝高的相关系数最大,在3个生态环境下相关系数均在0.900以上;茎粗除在嵩明点与小叶宽、在巧家点与叶长宽比和小叶宽相关不显著外,与其他农艺性状均呈极显著正相关;第一分枝高除在峨山点与小叶宽、在巧家点与叶宽、叶长宽比、小叶宽相关不显著以及在嵩明点与小叶宽呈显著负相关外,与其他农艺性状均呈显著或极显著正相关;分枝数除在峨山点与小叶宽、在巧家点与叶宽、叶长宽比相关不显著以及在嵩明点和巧家点与小叶宽呈极显著负相关外,与其他农艺性状均呈显著或极显著正相关。鲜重在有数据的嵩明点和巧家点与株高、茎粗、第一分枝高、分枝数、分枝长、叶长6个农艺性状间均呈极显著正相关关系。而叶长宽比、小叶宽在3个生态环境下与某些农艺性状呈负相关。
主要农艺性状间相关性分析表明,不同生态环境下大麻主要农艺性状相关性存在差异,如分枝数与叶长在3个生态环境下都表现出正相关性,但显著性存在差异。在巧家地区种植的F2代群体中2个性状相关性达到显著水平,相关系数为0.164,而在峨山和嵩明地区种植的F2代群体中2个性状相关性则达到了极显著水平,相关系数分别为0.687和0.446。
表4 峨山地区F2代群体中9个农艺性状相关性分析

注:**表示在0.01水平(双尾)上相关性显著,*表示在0.05水平上相关性显著。下同
表5 嵩明地区F2代群体中10个农艺性状相关性分析


表6 巧家地区F2代群体中10个农艺性状相关性分析

2.4 F2代农艺性状遗传力分析
对10个农艺性状进行频率分布统计,结果表明10个农艺性状频率分布均表现出单峰分布,偏度和峰度绝对值在大部分生态环境下小于1(表3),且具有连续性较好的正态分布趋势(图1为4个农艺性状的频率分布图),表明10个农艺性状属多基因控制的数量性状,但也有部分农艺性状在不同的生态环境下表现出正偏性,如第一分枝高在峨山地区种植下,其偏度绝对值大于1,表现出明显正偏性,而在另外两个生态环境下偏度和峰度绝对值均小于1,表明不同的生态环境会对部分农艺性状产生较大影响。

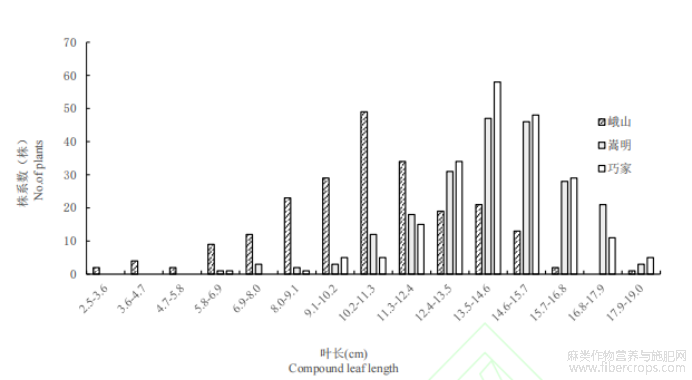
图1 3种生态环境下4个农艺性状频率分布图
将10个农艺性状进行表型方差分析,结果表明,10个农艺性状同时受到基因型、环境、基因型与环境互作(G×E)的极显著影响(P<0.01)。估算出10个农艺性状广义遗传力(表7),10个农艺性状广义遗传力由大到小依次为分枝长>株高>第一分枝高>叶长>分枝数>茎粗>叶宽>小叶宽>鲜重>叶长宽比,其中前5个农艺性状广义遗传力均超过65%,表明这5个农艺性状主要受遗传影响,具有较高的遗传效应,广义遗传力最高的是分枝长和株高,分别为82.575%和81.811%。而广义遗传力最小的农艺性状是叶长宽比,仅为29.508%,表明该性状遗传效应较小,主要受环境影响,由亲本难以预测子代的表型。
表7 10个农艺性状方差组成及广义遗传力估算
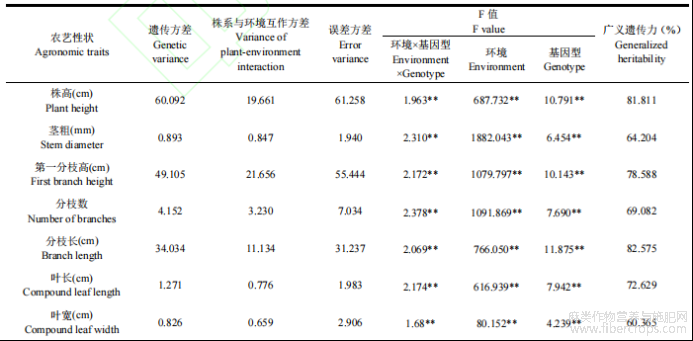

注:**表示在0.01水平上差异极显著。
3讨论
3.1 F2代群体中丰富的遗传变异可为大麻品种选育提供材料来源
中国大麻种质资源丰富,种植历史可追溯至5千年以上[19],并在长期的种植过程中形成了具有地方特色的种质资源,这些种质资源大多集中在籽用、纤维用和籽纤兼用等方面,且主要属于sativa生态型,普遍存在植株高大、分枝稀疏、叶片大等特征,显著区别于原产印度等南亚地区的indica生态型。随着工业大麻多用途和机械化的开发应用,尤其是药用工业大麻的兴起,矮秆、早熟、多分枝、富含大麻素等农艺性状备受关注。杂交群体农艺性状的多样性水平和变异程度对于培育作物新品种具有重要意义[20]。因此,本研究选取两份来自不同生态型且遗传背景差异较大的大麻材料作为亲本,并成功构建F2代分离群体,对F2代分离群体中10个主要农艺性状进行评价,结果表明10个农艺性状分离广泛,变异系数均大于10%,其中第一分枝高和鲜重变异系数较大,与汤志成等[14]通过对16份大麻种质资源进行表型变异分析,得出第一分枝高和地上生物量变异程度较大的结果一致。在本研究中,F2代分离群体的性状变异相比自然变异更加丰富,表明利用大麻不同生态型材料进行杂交,可以在分离后代中获得丰富的表型变异单株,这为不同用途的大麻品种选育提供了宝贵的材料来源。
3.2不同生态环境对大麻植株农艺性状的影响
尽管大麻分枝长、株高等性状遗传力较高,主要受遗传控制,但不同的生态环境仍然对各农艺性状产生较大影响,如峨山地区种植的F2代株系群体中表现出株型矮小、分枝数少、分枝较短等特点,而在嵩明和巧家地区种植的F2代株系群体则株型相对高大、第一分枝明显增高、分枝数显著增多。部分农艺性状对环境敏感可理解为是对环境的适应,但在实际栽培中要特别注意对环境比较敏感的农艺性状,这部分农艺性状往往决定着品种的适应范围。如适合在北方种植的大麻引种至南方则会提前开花,生物量降低[21]。而南方大麻移栽至北方则往往表现为迟熟,株型高大,生物量增加[22]。
不同生态环境对F2代株系群体中部分农艺性状产生显著影响的原因可能在于土壤、海拔和气候等环境因素不同而导致,尤其是光照对大麻生长发育的影响,因大麻是短日照植物,对光周期特别敏感[23],而大麻生物量主要由营养生长期积累,当光照时长低于开花所需临界时长大麻植株会进入生殖生长[24],导致生物量积累速度放缓,株型变得矮小。本研究中,虽然同一生态环境下F2代分离群体开花时间相对一致,但在不同纬度的3个环境下开花时间还是产生了差异,开花时间顺序呈现出峨山略早于嵩明,嵩明略早于巧家,导致除叶长外各农艺性状在3个生态环境下均产生显著差异。因此,在大麻育种和栽培过程中,应特别注意环境对大麻农艺性状和生物产量的影响,并为不同用途和栽培环境选育合理的株型品种。同时也可通过改良栽培措施,促进某些性状的发育,从而达到增产的目的。
3.3性状相关性分析和广义遗传力估算对农艺性状选择的影响
通过分析各农艺性状间关系,可以根据育种目的合理协调各农艺性状的关系,获得理想株型,提高育种效率。本研究相关性分析表明10个农艺性状间关系密切,除小叶宽、叶宽、叶长宽比3个性状外,其余农艺性状间均两两呈显著或极显著正相关关系。其中,株高与茎粗、叶长、叶宽都存在正相关关系,这与房郁妍等[25]对13份大麻种质材料农艺性状相关性研究结果一致。各性状间关系密切提示在大麻育种过程中,应综合多个农艺性状进行分析,在对目标性状进行选择的同时要考虑到对其关联性状的影响。
遗传力是研究数量性状遗传差异的主要参数之一,遗传力的大小可以为农艺性状世代选择提供参考[26],在本研究构建的F2代株系群体中株高和茎粗的广义遗传力分别为81.811%和64.204%,这与Petitd等[16]在3个生态环境下对123份大麻种质资源进行评价,得出株高和茎粗广义遗传力分别为0.81和0.69的结果基本一致。通过对各农艺性状进行广义遗传力估算,结果表明分枝长、株高、第一分枝高、叶长和分枝数5个性状广义遗传力均超过65%,表明这5个性状主要由遗传因素决定,在品种选育时可以从早期世代进行选择。而叶长宽比和鲜重广义遗传力则较低,表明这2个性状易受到环境的影响,早期世代不宜对这两个性状进行选择。
4结论
选用2份来自不同生态型且遗传背景差异较大的大麻材料作为亲本,成功构建F2代全雌性株系群体。对3种生态环境下F2代株系群体中10个农艺性状进行遗传变异及相关性分析。结果表明:8个农艺性状在3种生态环境种植下表现出显著差异(P<0.05);10个农艺性状表现出广泛的性状分离,为大麻不同用途品种选育创造了丰富的遗传变异类型;株高、分枝数、分枝长等性状频率分布具有典型的数量遗传特征,可通过QTL对相关基因进行定位;相关性分析表明10个农艺性状关系密切,提示在育种过程中突出单个性状的同时要考虑到对其关联性状的影响;分枝长、株高、第一分枝高、叶长及分枝数的广义遗传力均超过65%,其中分枝长和株高广义遗传力超过80%,可为大麻育种过程中农艺性状世代选择提供理论参考。以上结果可为大麻不同用途品种选育和相关性状的QTL定位提供一定的理论基础。
参考文献
[1]郭鸿彦.旱地工业大麻高产优质栽培技术[M].昆明:云南民族出版社,2013.
[2]Small E. Evolution and classification of Cannabis sativa (Marijuana, Hemp) in relation to human utilization[J]. Botanical Review,2015,81(3):189–294.
[3]SalentijnE M J, Zhang Q Y, Amaducci S, et al. New developments in fiber hemp (Cannabis sativa L.) breeding[J]. Industrial Crops and Products,2015,68(2):32–41.
[4]熊和平.麻类作物育种学[M].北京:中国农业科学技术出版社,2008.
[5]陈璇,郭蓉,万睿璇,等.云南省5个雌雄异株工业大麻群体遗传结构评价[J].分子植物育种,2015,13(9):2069–2075.
[6]邓纲,郭鸿彦,顿昊阳,等.环境因子对大麻纤维产量和质量影响的研究进展[J].中国麻业科学,2010,32(3):176–182.
[7]刘浩.影响工业大麻产量因素的研究[D].昆明:云南大学,2015.
[8]唐慧娟,臧巩固,程超华,等.工业大麻产量和品质性状的对应分析[J].作物杂志,2018(2):52–55.
[9]张晓艳,孙宇峰,曹焜,等.纤用工业大麻雌雄株主要农艺性状的初步研究[J].中国农学通报,2020,36(20):1–6.
[10]刘振业,刘貞琦.作物的株型与产量-作物高产的基础理论(三)[J].耕作与栽培,1982(6):58–65.
[11]王庆峰,张雪,李庆鹏,等.工业大麻种质资源农艺性状初步评价[J].农业与技术,2020,40(10):34–38.
[12]陈璇,杨明,郭鸿彦.大麻植物中大麻素成分研究进展[J].植物学报,2011,46(2):197–205.
[13]Josefina F, Robert V. PKS activities and biosynthesis of cannabinoids and flavonoids in Cannabis sativa L. plants[J]. Plant Cell Physiology,2008,49(12):1767–1782.
[14]汤志成,陈璇,张庆滢,等.野生大麻种质资源表型及其RAPD遗传多样性分析[J].西部林业科学,2013,42(3):61–66.
[15]吕咏梅.大麻品种主要农艺性状的遗传初探[J].中国麻作,1983(2):44–48.
[16]Petit J, Salentijn E M J, Paulo M J, et al. Genetic variability of morphological, flowering, and biomass quality traits in hemp (Cannabis sativa L.)[J]. Frontiers in Plant Science,2020,11:1–17.
[17]粟建光,戴志刚.大麻种质资源描述规范和数据标准[M].北京:中国农业出版社,2006.
[18]李辉.马铃薯高世代无性系矿质元素的稳定性及广义遗传力分析[D].哈尔滨:东北农业大学,2019.
[19]孙永刚.大麻栽培起源与利用方式的考古学探索[J].农业考古,2016(1):16–20.
[20]杨彦伶,李振芳,徐红梅,等.紫薇与福建紫薇种间杂交F1代遗传性状分析[J].东北林业大学学报,2018,46(9):31–34.
[21]姚青菊,熊豫宁,彭峰,等.不同生态类型大麻品种在南京引种的生育表现[J].中国麻业科学,2007,29(5):270–275.
[22]胡学礼,郭鸿彦,刘旭云,等.云南工业大麻品种在黑龙江大兴安岭地区的适应性研究[J].西南农业学报,2012,25(3):838–841
[23]孙宇峰.纤维大麻高产栽培技术的研究现状[J].中国麻业科学,2017,39(3):153–158.
[24]Amaducci S, Colauzzi M, Bellocchi G, et al. Evaluation of a phenological model for strategic decisions for hemp (Cannabis sativa L.)biomass production across European sites[J]. Industrial Crops and Products,2012,37(1):100–110.
[25]房郁妍,张利国,郑楠,等.大麻种质资源农艺性状的聚类分析、相关性分析及主成分分析[J].黑龙江农业科学,2018(10):18–21.
[26]余本勋,顾尚敬,潘亚昌.贵州高原粳稻主要经济性状遗传力和遗传进度的研究[J].贵州农业科学,1992(6):13–16.
文章摘自:张坤,陈璇,杜光辉,郭鸿彦,郭蓉,吕品,郭孟璧,许艳萍,张庆滢,杨明.两种生态型大麻杂交F2代主要农艺性状遗传变异分析[J/OL].西南农业学报.
